dead ash along Accotink Creek, Fairfax County, Virginia photo by F.T. Campbell
As we all know, the emerald ash borer (EAB) has killed millions of ash trees in its invaded range across eastern North America. However, field studies have detected robust regeneration of ash seedlings and saplings in various invaded areas. Ward et al. 2021 (full citation at end of blog) set out to determine whether this regeneration will result in recovery of mature ashes that can perform their ecological role. They conclude that it will not. Instead, they say, the EAB invasion will probably alter successional patterns and composition of large areas of naturally regenerating forests, causing a cascade of ecological impacts in ash-containing ecosystems
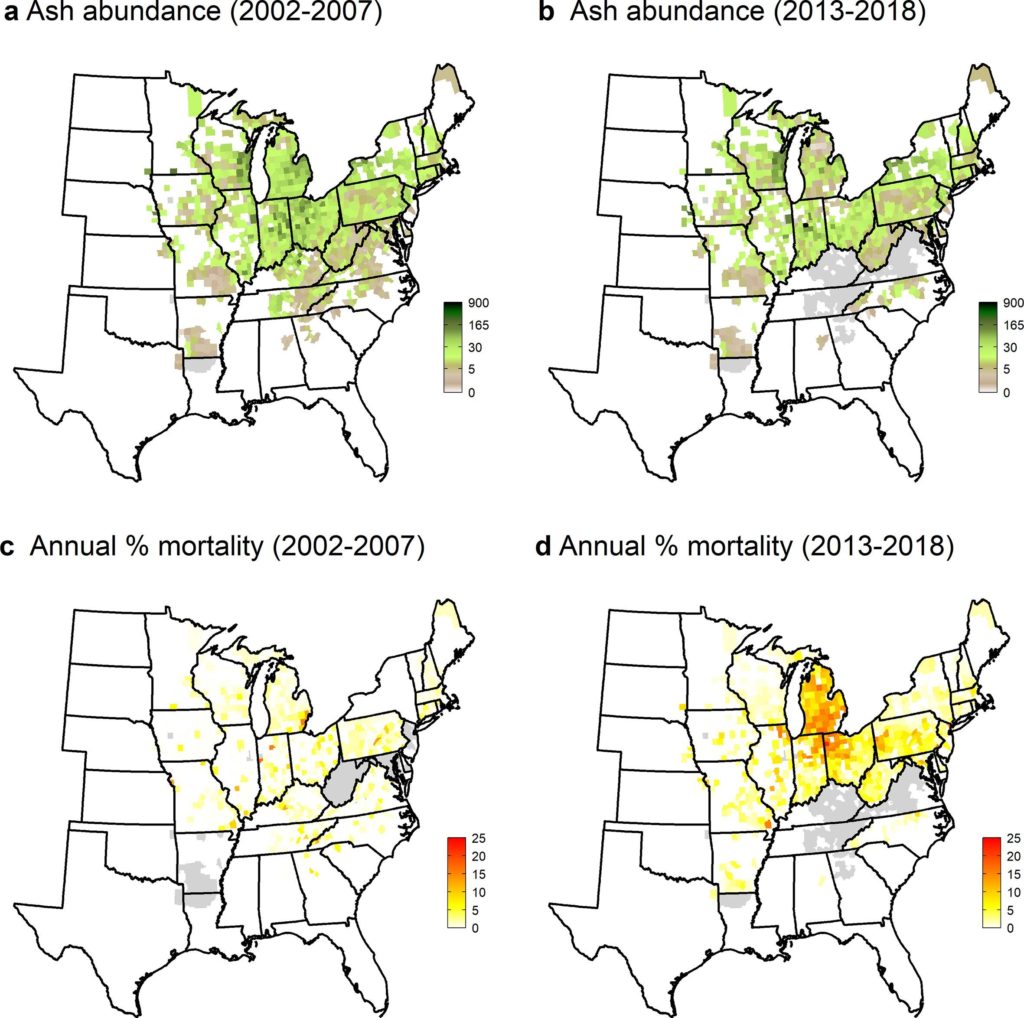
Ward and colleagues used USDA Forest Service Forest Inventory and Analysis (FIA data) to quantify ash recruitment and regeneration across the entire eastern United States. Theirs is the first study to evaluate trends across the region, rather than specific locations or stands. They related the FIA recruitment data to EAB spread, as measured by USDA Animal and Plant Health Inspection Service’ (APHIS) record of the first EAB detection in each county.
FIA inventories in 2002-2007 and 2013-2018 show large numbers of ash seedlings and saplings in counties invaded in the first wave of invasion, 2002–2006. These areas had higher densities of both seedlings and saplings than plots in other counties. The earliest-invaded counties were in areas that had extraordinarily high densities of ash before the EAB invasion, so the numbers of seedlings and saplings probably reflected that abundant seed source.
However, by the 2013-2018 inventory ash trees in the smallest overstory class (12.7 cm dbh) were dying at faster rates than they were recruited from seedlings or saplings in all 362 counties recorded by APHIS as EAB-infested before 2013. Ward and colleagues found these negative population trajectories on plots that have been invaded for more than about 10 years. This trend suggests that ash will continue to decline in abundance and may become functionally extinct across the invaded range.
Some U.S. Forest Service biologists are more optimistic about ash recovery in response to biocontrol of the EAB. See their podcast here.
In the risk of functional extinction, ash trees are unfortunately not unique. The authors note similar impacts from the invasion of the hemlock woolly adelgid and beech bark disease.
Data Reveal History of Invasion (spread)
Ward and colleagues focused on the risk of mortality for young ashes as they developed from seedlings to saplings, and, eventually, to overstory trees. The youngest “overstory” trees are 12.7 to 17 cm dbh. FIA data show that even the largest trees in this class are 3 cm smaller than trees that produce seeds.
Mortality was initially uniformly low – less than 2.1% — as measured by the first FIA inventory (2002–2007). This is not surprising because EAB was detected only in 2002, and then in only few counties. (EAB had probably been present for a decade before it was detected.)
By the 2013-2018 FIA inventory, mortality had quadrupled to 8–11% in counties invaded during the 2002–2006 period. In the counties invaded during the 2007–2012 period, morality also rose to 3-5%. Both measurements included all diameter classes. Annual mortality rates in the FIA 2013-2018 inventory were still highest for the counties invaded during 2002–2006 except for the largest trees (those greater than 40 cm dbh). By the time of the 2013-2018 FIA survey, overstory ash densities near the epicenter had since declined substantially. They had been nearly eliminated in some counties in southeastern Michigan. There were still sufficient numbers of smaller trees in the region to exhibit an elevated mortality rate – more than 10% per year in several counties in Michigan, Indian, and Ohio. By contrast, in the most recently invaded areas – those counties recorded by APHIS as infested after 2013 – there was very little change in ash densities compared to the 2002-2007 period. This is hardly surprising since it takes years for mortality to reach levels observable by the FIA process.
dead ash on edge of Pohick Bay, Fairfax County, Virginia photo by F.T. Campbell
Considering trees just entering the overstory category (those with diameters of 12.7 cm dbh), annual mortality increased substantially across the region. Between the first FIA inventory (conducted in 2002-2007) and the second inventory (conducted in 2013-2018), their average annual mortality rose more than four-fold, from 0.08 trees per ha to 0.37 trees per ha. By 2013-2018, recruitment in the 2002–2006 invasion cohort was about 50% less than tree mortality levels; recruitment and mortality were about equivalent for the counties invaded in the 2007–2012 period. Recruitment was [still] significantly higher than mortality for the counties recorded as invaded in 2013–2018. However, Ward and colleagues expect mortality rates of this cohort to accelerate over the next five to 10 years – even in areas with lower ash densities.
Ward and colleagues note that many of the young ash trees were dying before they could reach reproductive age – which they estimated to be about 20 years with a dbh of about 20 cm.
As the invasion progresses and hosts are depleted, mortality rates could slow, but, for ash to persist, it is critical that sufficient numbers of trees reach reproductive age before succumbing to residual EAB populations.
Other factors that might influence ash include competition with trees in other genera. The biocontrol agents now becoming established in young ash forests might increase the likelihood of ash persistence. Still, seed production and seedling survival will need to be frequent and widespread if they are to offset expected mortality. Resilience might also vary depending on individual species’ vulnerability to changes in the climate and to EAB (green and black ash are more vulnerable than white ash).
SOURCE
Ward, S.F., A.M. Liebhold, R.S. Morin, S. Fei. 2021. Population dynamics of ash across the eastern USA following invasion by emerald ash borer. Forest Ecology and Management 479 (2021) 118574
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
eastern hemlocks in Great Smoky Mountains National Park
As
we all know, eastern (Tsuga canadensis)
and Carolina (T. caroliniana) hemlocks
have suffered huge losses due primarily to the introduced hemlock woolly
adelgid (Adelges tsugae – HWA). In New England, there has been more
than a 60% decrease in total hemlock basal area since 1997 and a virtual
absence of hemlock regeneration in HWA-infested areas. HWA continues to spread
– most recently into western Michigan and Nova Scotia (all information, unless
otherwise indicated, is from Kinahan et
al. 2020; full citation at end of this blog). [However, Morin and Liebhold
(2015) found that hemlock basal volume continued to increase for the first 20
years or so after invasion by the adelgid, due to ingrowth of immature
hemlocks. See “results” in Morin et al.,
full citation at the end of the blog.]
This
loss deprives us of a gorgeous tree … and unique habitats. Hemlock-dominated
forests were characterized by deep shade, acidic and slowly decomposing soil,
and a cool microclimate. They provided unique and critical habitat for many
terrestrial and aquatic species.
A
team of scientists based at the University of Rhode Island has carried out an
experiment comparing cuttings from eastern hemlocks apparently resistant to HWA
to susceptible ones. Matching sets of resistant and susceptible trees were
planted at eight sites in seven states – Ithaca and Bronx, NY; Boston; southern
CT; Lycoming County, PA; Thurmont, MD; southern WV; and Waynesville, NC. All plantings
were within or adjacent to forests containing HWA-infested hemlocks.
After
four years, 96% of the HWA-resistant hemlocks had survived, compared to 48% of
the control plants. The HWA-resistant plants were 32% taller, put out 18% more
lateral growth, had 20% longer drip lines, and were in 58% better condition.
HWA was found on trees at only three out of the eight plots. HWA density on
resistant eastern hemlocks was 35% lower than on HWA-susceptible hemlocks,
although this difference was not statistically significant.
Trees
in all eight plots were infested with elongate hemlock scale (Fiorinia externa – EHS), a second insect
damaging hemlocks in eastern North America. However, the HWA-resistant hemlocks
had EHS densities 60% lower than those of the controls.
Kinahan et al. note that identification and use of host tree populations’ potential for pest resistance has played a role in other programs managing non-native pests and pathogens, including Dutch elm disease and chestnut blight.
The
same scientists note that significant effort has been put into biocontrol or
insecticides for management of hemlock woolly adelgid, but without achieving the
desired improvement of forest health. Attempts to cross eastern hemlocks with
HWA-resistant hemlocks unfortunately produced no viable offspring. However,
Kinahan et al. were inspired to
explore possible genetic resistance within natural populations of eastern
hemlocks by the 1) evidence of resistance in Asian and western hemlocks; 2) the
different foliar terpene profiles in those species; and 3) the presence of
apparently healthy mature hemlock trees growing in proximity to heavily
infested trees.
They asked forest managers and other concerned groups to help locate stands with trees that were mature and apparently completely healthy, were located within HWA-devastated hemlock stands, and had not been chemically treated. They chose a small stand of eastern hemlocks growing within the Walpack Fish and Wildlife Management Area in northern New Jersey. This stand was called the “Bulletproof Stand”. They evaluated HWA resistance in five of these trees, then chose two for propagation and planting in the test.
New Jersey’s “bullet-proof stand” on the left photo by Richard Casagrande
The
trees were planted in September 2015. Due to funding gaps, they were not
revisited for four years. Thus, Kinahan et
al. re-evaluated the resistant and vulnerable trees in Autumn 2019 – with
the results I reported above.
Does this study
prove that clonal propagation of apparently resistant hemlocks is an effective
strategy to restore the species?
It
is not that simple.
The
difference in survival and condition was striking, but the authors note several
caveats:
1)
they had not recorded pre-experiment data on plant height or other variables,
so they cannot be certain that variation in initial plant height or dripline
did not contribute to current treatment-level differences in these variables.
2)
they cannot distinguish between the impacts of HWA and EHS on plant growth.
3)
since they could not monitor the planting sites for four years, they cannot
definitively link increased mortality of HWA-susceptible trees to higher pest
densities. However, the lower pest densities and higher survival of
HWA-resistant hemlocks are consistent with herbivore-driven tree mortality.
They
also cannot assess the impact of other environmental stressors (drought, cold,
etc.) on their results.
4)
The small number of trees planted at each site prevented detailed site-level
analyses.
The
scientists conclude that their work is most appropriately viewed as a ‘proof of
concept’ experiment highlighting the need
for future research exploring how HWA-resistant eastern hemlocks might best be
integrated into existing HWA management.
Unfortunately, the
Rhode Island researchers report they cannot persuade the US Forest Service to
support continuing this effort. Will these promising hints not result in
action?
Kinahan
et al. stress the importance of the
reduced pest densities (both HWA and EHS) on the putatively resistant hemlocks.
They think this might be a result of the higher terpene concentrations in the
twigs and needles. Finally, they note that lower densities of sap-feeding
herbivores may also indirectly provide protection against other consumers,
including gypsy moth (Lymantria dispar)
and hemlock looper (Lambdina fiscellaria).
SOURCE
Kinahan,
I.G., G. Grandstaff, A. Russell, C.M. Rigsby, R.A. Casagrande, and E. L.
Preisser. 2020. A four-year, seven-state reforestation trial with eastern hemlocks
(Tsuga canadensis) resistant to hemlock woolly adelgid (Adelges tsugae).
Forests 11: 312
Morin, R.S. and A.M. Liebhold. 2015. Invasions by two non-native insects alter regional forest species composition and successional trajectories. Forest Ecology and Management 341 (2015).
Posted by Faith Campbell
P.S. I have been working with colleagues to promote a more coordinated and well-funded program to combat non-native forest pests – including much greater reliance on identifying and breeding resistance to the pest. Visit here to see this effort.
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
redbay in Georgia killed by laurel wilt photo by Scott Cameron
In August I posted a blog summarizing information on pest introductions and impacts gleaned over my nearly 30 years of engagement. Already, I need to post an update, with an alarming estimate of introduced pests’ impacts across the continent.
Fei
et al. 2019 (see the full citation is
at the end of this blog) estimate that the
15 most damaging introduced species threaten 41.1% of the total live forest
biomass in the 48 conterminous states.
In
fact, this might be an underestimate
of the pests’ impacts on biomass loss. Fei et
al. (2019) note several limitations in their data that might result in such
an underestimate. These include:
1)
Mortality rates – and impacts – may increase over several decades following the
initial invasion.
2)
For pests already established throughout nearly all their potential ranges,
pest-induced biomass loss could be substantially underestimated because most of
their hosts died long ago, before the FIA data began to be collected. Consequently,
the actual loss of these tree species from the forest is much greater than has
been measured by the study’s methods.
3)
Mortality rates vary among species and regions, which might introduce errors.
They cite European gypsy moth, in which relatively small areas of heightened
mortality due to repeated defoliations are swamped by lower mortality rates
across the chosen measurement area.
4)
They considered only tree mortality, not crown or root dieback or reductions in
tree growth.
5)
They did not estimate carbon release to the forest floor as a result of
defoliation.
6)
Pest-related mortality rates may be underestimated due to salvage – although
the authors did not observe evidence of substantial salvage efforts for most of
these pests.
7)
The data did not include losses from urban tree mortality.
Fei
et al. estimate that more than 450 pests
are established in the 48 conterminous states. This study excluded pests
attacking palms; trees on U.S. Pacific and Caribbean islands; and pests native
to portions of North America that are causing severe damage in naïve hosts – e.g., goldspotted oak borer. I did
include the latter groups in my slightly larger estimate laid out in my August
blog.
Fei
et al. base their analysis on 83 of
the introduced pest species considered to cause substantial effects on tree
health and productivity and sometimes extensive tree mortality. (In my August
blog, I described findings by another study by Guo et al (2019) that counted 91 species in that category.)
Fei et al. build on studies by a group of USDA Forest Service (USFS) scientists that I described in an earlier blog.
This team found that, nation-wide, non-native forest pests are causing an approximate 5% increase in total annual mortality by tree volume. They based their studies on analysis of 92,978 long-term plots maintained by the USDA Forest Inventory and Analysis program.
As
noted above, the article cannot capture the full range of mortality in species
affected by pests introduced decades ago. Chestnut blight, white pine blister
rust, Port-Orford-Cedar root disease, beech bark disease, butternut canker,
dogwood anthracnose, and European gypsy moths had all killed millions of trees
before the USFS forest inventory plots were established. Fei et al. do form a solid basis for measuring some of the current
impacts and projecting future ones.
The
focus of the new article is on the amount of carbon being transferred from live
biomass to dead organic matter as a result of the increased mortality caused by
the 15 species with the highest impacts. This is arguably a more quantifiable
measure of pests’ impacts than others’ approaches. Here, I focus more narrowly on the
documentation of exacerbated mortality as measured by the loss of biomass. Added
together, these 15 species have caused an additional (i.e., above background levels) tree mortality rate of 5.53 TgC per
year [defined as terragrams of carbon]. This
estimate of annual conversion of live biomass to dead wood is similar in
magnitude to that attributed to fire (5.4 to 14.2 TgC per year) (Fei et al. 2019). Yet the fire threat gets
much more attention – for both prevention and management.
It
is important to remember that conversion of living biomass to dead wood does
not result in an immediate release of carbon to the atmosphere. Atmospheric releases
take place through decomposition which is both gradual and takes place at
varying rates. Some of the carbon will remain in the soil. And, over time, some
of the carbon storage capacity will be restored by compensatory growth in
unaffected trees and the recruitment of new regeneration – although this faster
growth is delayed by as much as two or more decades after pest invasions begin
(Fei et al. 2019).
American elm being killed by “Dutch” elm disease photo by Cheryl Kaiser, University of Kentucky; bugwood.org
The 15 species of introduced pests used in this analysis
are laurel
wilt disease, chestnut blight, butternut canker, dogwood anthracnose, emerald
ash borer (EAB), Dutch elm disease, red pine scale, beech bark disease, hemlock
woolly adelgid, balsam woolly adelgid, European gypsy moth, white pine blister
rust, green spruce aphid, sudden oak death, and Port-Orford cedar root disease.
Of these, the highest elevation in biomass loss – as measured by FIA plot data
– was caused by EAB, Dutch elm disease, beech bark disease, and hemlock woolly
adelgid. We know that elms and beech, at least, began dying decades before the
FIA data began to be collected. So the reported mortality rates are an
underestimate. This is especially true because beech mortality is highest in
the first decade after invasion by beech bark disease.
Annual levels of biomass loss are virtually certain to increase. First, pests will spread to new host ranges and infestations in already-invaded ranges will intensify. As a result, substantial amounts of the hosts’ biomass are at risk of exacerbated mortality. As I noted at the top of the blog, the total amount of host biomass at risk from these 15 species is estimated to be 5,197 TgC – or 41.1% of the total live forest biomass in the 48 conterminous states. Further exacerbating future losses is the likelihood that additional pests will be introduced. I would add that pests not included in this analysis – e.g., polyphagous and Kuroshio shot hole borers and possibly the spotted lanternfly – are also likely to contribute to losses of live forest biomass.
Fei
et al. (2019) did not attempt to
determine the economic value of this biomass loss or to address other types of
losses to ecosystem services.
Remember that a separate set of studies reported by Potter et al. (2019) (the CAPTURE project) also relied on data from the FIA plots to evaluate the impact of introduced pests. These studies focused on identifying the host species at greatest risk rather than highest-impact pests or biomass loss. I find it reassuring that the Fei and Potter studies – using different approaches – resulted in very similar species rankings. See my discussion of the Potter studies here.
Together, the teams led by Potter and Fei set clear priorities for addressing the threats from non-native pests. What we need now is action! See my recommendations in my recent “solutions” blog.
U.S. Capitol – inform your representatives that you want action to protect trees!
SOURCES
Fei,
S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from
insect and disease invasions in United States forests
Potter,
K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019a. Important Insect and
Disease Threats to United States Tree Species
and Geographic Patterns of Their Potential Impacts. Forests. 2019 10 304.
Potter,
K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019b. Prioritizing
the conservation needs of United States tree species: Evaluating vulnerability
to forest insect and disease threats. Global Ecology and Conservation. (2019)
Faith Campbell receives award for activism from National Association of State Foresters; 2016
For nearly 30 years I have documented bioinvasion threats and gaps, first in three Fading Forests reports (available here), then in five years of blogging. Here I pull together that information and suggest — in most cases reiterate — steps to address these threats and gaps. I list sources of discussion of the underlying issues – other than my reports and blogs – in references at the end of this blog.
My
first premise is: robust federal leadership is crucial:
The Constitution gives primacy to
federal agencies in managing imports and interstate trade.
Only a consistent approach can
protect trees (and other plants) from non-native pests.
Federal agencies have more
resources than state agencies individually or in any likely collective effort
— despite decades of budget and staffing cuts.
My
second premise is: success depends on a continuing, long-term effort founded on
institutional and financial commitments commensurate with the scale of the threat.
This requires stable funding; guidance by research and expert staff; and engagement
by non-governmental players and stakeholders. Unfortunately, as I discuss
below, funding has not been adequate or stable.
My third premise is that programs’ effectiveness needs to be measured, not just effort (see the NECIS document referenced at the end of the blog).
SPECIFICS
Preventing
new introductions continues to be the most effective action. Mitigating options
decrease and damages increase once a non-native pest has entered the country –
much less become established (see Lovett et
al. 2016 and Roy et al. 2014). I
recognize that preventing new introductions poses an extremely difficult
challenge given the volume and speed of international trade and the strong
economic forces supporting free trade. These challenges have been exacerbated over
several decades by the political zeitgeist – the anti-regulatory ideology, the emphasis
on “collaborating” with “clients” rather than imposing requirements through
regulations. Although the current “America First” policy might reduce import
volumes and therefore reduce the invasive species threat to some extent, the
anti-regulatory stance has only strengthened.
containers at the Port of Long Beach, California
Decades of cutting key agencies’ budgets and personnel are another factor. However, the damage to America’s natural systems is so great that we must try harder to find more effective strategies (See the Fading Forest reports; my previous blogs; Lovett et al. 2016; and APHIS annual reports – e.g., the 2019 report here)
Prevention
Despite adoption and implementation of new international and national regulations to stem pest introductions, introductions continue – although probably at a lower level than would otherwise be the case. Delays in adoption of regulations (documented in Fading Forests II and III and my two recent 30-years-in-review blogs have facilitated damaging introductions and spread.
Solutions
Stakeholders press USDA
leadership to initiate rules intended to strengthen phytosanitary protection and
expedite their completion
APHIS promote and facilitate
analysis of current programs and policies by non-agency experts to ensure the
agency is applying most effective strategies (Lovett et al. 2016).
Adoption
of insufficiently protective regulations (documented in FFII, FFIII, two
30-years-in-review blogs) – adopted in part because APHIS is trying to
“balance” trade facilitation and phytosanitary protection – has further
contributed to damaging pests’ introduction and spread.
Solutions:
Boost
priority of preventing pest introductions by amending the Congressional finding
in the Plant Protection Act [7 USC 7701(3)] as follows
Existing language: “[I]t is the
responsibility of the Secretary [of Agriculture] to facilitate exports, imports
and interstate commerce in . . . commodities that pose a risk of harboring
plant pests or noxious weeds in ways that
will reduce, to the extent practicable, as determined by the Secretary, the
risk of dissemination of plant pests and noxious weeds .… “
Amend to read as follows: “…. in ways that will reduce prevent, to the greatest extent practicable feasible, as determined by the Secretary, …” [emphasis added]
Adopt several actions to
strengthen phytosanitary protections at the point of origin (Lovett et al. 2016)
Expand
pre-clearance partnerships — as authorized for plants under Q-37 regulations
and ISPM-36
Expand
sentinel tree programs
Promote
voluntary substitution of packaging made from materials other than solid wood.
APHIS
doesn’t use the enforcement powers that it has under Plant Protection Act (see
several of my past blogs)
Solutions:
CBP inspectors search for pests in a pallet; CBP photo
APHIS follow the lead of Customs and Border Protection and begin penalizing importers on the first instance of their wood packaging not being in compliance with ISPM#15 (see blog here).
APHIS prohibit use of wood packaging by countries and importers of categories of imports that – over the 13 years since implementation – have developed a record of frequent violations of ISPM#15.
APHIS use its authority per revised Q-37 regulations to negotiate with countries that export plants to the U.S. to establish “integrated measures” programs aimed at minimizing the risk of associated pests being transported to the U.S.
APHIS use its authority per revised Q-37 to place in the “Not Authorized for Import Pending Pest Risk Assessment (NAPPRA) “limbo” category genera containing North American “woody” plants (see Roy et al. 2014; Lovett et al. 2016).
Spread within the
U.S.
The
United States lacks a coordinated system to prevent pest spread within the
country (see Fading Forests III Chapter 5). Even our strictest methods, like APHIS’s
quarantines regulating interstate movement of goods, have failed to curtail
spread of significant pests. The most obvious example is the emerald ash borer.
The regulations governing movement of the sudden oak death pathogen in the nursery trade have also failed: there have been periodic outbreaks in which the pathogen has been spread to nurseries across the country. Between 2003 and 2011, a total of 464 nurseries located in 27 states tested positive for the pathogen, the majority as a result of shipments traced from infested wholesalers. In 2019, plants exposed to the pathogen were again shipped to 18 states; eight of those states have confirmed that their plant retailers received infected plants (see my blog from summer here).
Another
serious gap is the frequent failure of APHIS and states to adopt official
programs targetting bioinvaders that will be difficult to control because of
biological characteristics or cryptic natures – even when severe impacts are
demonstrated. Recent examples include the laurel wilt disease complex, goldspotted
oak borer, polyphagous and Kuroshio shot hole borers and associated pathogens,
and even the spotted lanternfly (although the last has received significant
funds from APHIS.)
redbay killed by laurel wilt disease, Georgia; photo by Scott Cameron
Solutions:
APHIS apply much more stringent
regulations to interstate movement, based on a heightened priority for
prevention in contrast to facilitating interstate trade. E.g., prohibit nurseries on the West Coast from shipping P. ramorum hosts to states where the pathogen
is not established.
APHIS encourage states to adopt
quarantines and regulations aimed at preventing spread of invasive pests to
regions of the state that are not yet infested. For example, the sudden oak
death pathogen in California and Oregon; the borers in southern California.
APHIS abandon plans to deregulate
emerald ash borer and step up its support for state regulations on firewood.
APHIS stop dumping pests it no
longer wants to regulate onto the states through the “Federally Recognized
State Manage Phytosanitary (FRSMP) program”.
APHIS revise its policies so that
the “special needs exemption” [7 U.S.C. 7756] actually allows states to adopt
more stringent regulations to prevent introduction of APHIS-designated
quarantine pests (see Fading Forests III Chapter 3).
To help fill the gaps, the states are trying to coordinate their regulations in some important areas. The most advanced example is the voluntary Systems Approach to Nursery Certification, or SANC program. APHIS has supported this initiative, including by funding from the Plant Pest and Disease Management and Disaster Program (see below). However, it is a slow process; USDA funds first became available in 2010. The states are trying to coordinate on firewood, but we don’t yet know what the process will be.
Funding shortfalls (See the three Fading Forests
reports, my blogs about appropriations)
Increase APHIS’ access to emergency
funds from the Commodity Credit Corporation by
amending the Plant Protection Act [7 U.S.C. 7772 (a)] to include this
new definition of “emergency”:
the term “emergency” means any
outbreak of a plant pest or noxious weed which directly or indirectly threatens
any segment of the agricultural production of the United States and for which
the then available appropriated funds are determined by the Secretary to be
insufficient to timely achieve the arrest, control, eradication, or prevention
of the spread of such plant pest or noxious weed.
Although APHIS has the most
robust prevention program of any federal agency, its funding is still
inadequate. Stakeholders should lobby the Congress in support of higher annual
appropriations.
The Plant
Pest and Disease Management and Disaster Program (now under Section 7721 of the
Plant Protection Act) has provided at least $77 million for tree-pest
programs (excluding NORS-DUC & sentinel plant programs and other programs)
since FY 2008. Much useful work has been carried out with these funds. However,
these short-term grants cannot substitute for stable, long-term funding. I
reiterate my call for stakeholders to lobby the Congress to provide larger
appropriations to the APHIS Plant Protection program and Forest Service Forest
Health Protection and Research programs.
Long-term Responses
to Bioinvasive Challenge
More stakeholders are advocating raising the priority of – and providing adequate resources to – such long-term solutions as biocontrol and breeding trees resistant to pests and restoring them to our forests. Advocates include the state forestry agencies of the Northeast and Midwest, some non-governmental organizations, some academics, and individual USFS scientists. One effort resulted in inclusion of language in the 2018 Farm Bill (see blog here) – although this approach has apparently run into a dead end. The new emphasis on breeding has so far not been supported by agency or Congressional leaderships.
test planting of an American chestnut bred to be resistant to chestnut blight
Solutions:
USFS convene workshop of the
federal, state, National Academy, academic, and NGO groups promoting resistance
breeding in order to develop consensus on priorities and general structure of program.
Explicitly include evaluation of the
CAPTURE Project’s (see blog here) efforts to
set priorities to guide funding allocations and policies; and proposals for
providing needed supportive infrastructure – facilities, trained staff in
various disciplines. (See my blogs here.)
Report results of meeting to USDA
leadership, Congress, and stakeholders
Then ensure implementation of the
accepted approach by both Research and Development and Forest Health Protection
programs. Include provisions to provide sustainable funding.
These proposed actions still do not address ways to correct the provisions of the international phytosanitary agreements (World Trade Organization and International Plant Protection Convention) that complicate – or preclude – efforts to prevent introduction of pests currently unknown to science. This issue is discussed in Fading Forests II. A current example is beech leaf disease (described here).
Continuing
inadequate engagement by stakeholders
Most
constituencies that Americans expect to protect our forests don’t press
decision-makers to fix the problems I have identified above: inadequate
resources, weak and tardy phytosanitary measures. Some of these stakeholders
are other federal agencies, or state agencies – or their staffs. They face
restrictions on how “political” they can be. But where are the professional and scientific associations,
representatives of the wood products industry, forest landowners, environmental NGOs and their funders, urban
tree advocates Efforts by me, USDA, and others to better engage these groups
have had disappointing results.
As
I have documented, groups of USFS scientists have made several attempts to
document the extent of invasive species threats and impacts and to set
priorities. So far, they have not gained much traction. Another USFS attempt,
Poland et al. in press, will appear
at the end of the year. Will this be more successful?
I
detect growing attention to educating citizen scientists for early detection;
but if there is an inadequate – or no – official response to their efforts
won’t people become discouraged?
SOURCES
Lovett,
G.M., M. Weiss, A.M. Liebhold, T.P. Holmes,
B. Leung, K.F. Lambert, D.A. Orwig, F.T. Campbell, J. Rosenthal, D.G. McCullough,
R. Wildova, M.P. Ayres, C.D. Canham, D.R. Foster, SL. Ladeau, and T. Weldy.
2016. NIS forest insects and pathogens in the US: Impacts and policy options. Ecological
Applications, 26(5), 2016, pp. 1437–1455
National Environmental Coalition on Invasive Species “Tackling the Challenge.”
Poland,
T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019),
Invasive Species in Forests and Grasslands of the United States: A
Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. (in press).
Roy,
B.A., H.M Alexander, J. Davidson, F.T Campbell, J.J Burdon, R. Sniezko, and C.
Brasier. 2014. Increasing forest loss worldwide from P&Ps requires new
trade regulations. Front Ecol Environ 2014; 12(8): 457–465
dead whitebark pine in Crater Lake National Park photo by F.T. Campbell
I began studying and writing about the threat to North America’s forests from non-native insects and pathogens in the early 1990s – nearly 30 years ago. I reported my analyses of the evolving threat in the three “Fading Forests” reports – coauthored by Scott Schlarbaum – in 1994, 2003, and 2014. These reports are available here.
So what has changed over those 30 years? What remains the same? Why have both the changes and the stasis occurred? What can we do to fix the gaps, close unaddressed pathways, strengthen flabby policies? I will address these issues in this and following blogs.
experimental American chestnut planted in Fairfax County, VA photo by F.T. Campbell
What has changed
since the early 1990s:
Adoption and implementation of significant new international and national regulations and programs aimed at preventing introductions of non-native invasive species.
Despite the welter of new regulations, an alarming increase in numbers of highly damaging forest pests established in the country. (By my count, about 50 new species have established on the continent, six on Pacific islands; see details below.)
Alarming spread of established pests to new geographic regions and new hosts (e.g., emerald ash borer in 35 states and 5 provinces; laurel wilt disease across the range of redbay and swamp bay; rapid ‘ōhi‘a death on three of the main Hawaiian islands).
Introductions via unexpected pathways and vectors far removed from phytosanitary agencies’ usual targets, e.g., ship superstructures, imported steel and stone …
What has remained the
same since the early 1990s:
Inadequate
resources provided to response and recovery efforts.
Available
funding focused on only a few of the more than 90 species causing damage.
Adoption
of insufficiently protective regulations that have failed to prevent
introduction and spread of tree-killing pests.
Lengthy
delays in implementing programs that tighten controls – another factor in
continuing introductions and spread.
Continued
importance of expected pathways – nursery stock and raw wood, especially
crates, pallets, and other forms of wood packaging.
Federal
and state agencies still choose not to take action on pests e.g., goldspotted oak borer, polyphagous
and Kuroshio shothole borers, beech leaf disease.
Inadequate
coordination despite several efforts to set priorities.
Spurts
of attention by media and political decision-makers, contrasted by lengthy
periods of inattention.
Failure
of most stakeholders to support efforts to prevent and respond to introductions
of tree-killing pests.
Details: The Situations
Then and Now
(Many of the individual species mentioned here are described more fully here. Full citations of sources are at the end of blog.)
American elm on the National Mall, Washington, D.C.
photo by USDA Agricultural Research Service
In 1993:
The number of non-native forest pest species established in the U.S. was estimated at between 300 (Millers et al. 1993) and 380 (Mattson et al., 1994; Liebhold et al., 1995) .
The area suffering the greatest numbers and impacts was the Northeast.
Several highly damaging pests that had been established for decades, including chestnut blight, white pine blister rust, Port-Orford-cedar root disease, Dutch elm disease, hemlock woolly adelgid, butternut canker, and dogwood anthracnose were receiving some attention but continued to spread.
USDA Forest Service funding for management of exotic pest infestations was crisis-oriented, with “… priorities … set under political pressures for immediate answers, with too much regard for short-term problems and too little consideration for broader management objectives.” (NAS 1975)
Since few high-profile pests had been introduced in recent years, APHIS was not actively engaged. In FY92, APHIS spent $20 million on efforts to eradicate the Asian gypsy moth. The narrow focus is illustrated by the fact that in FY93, more than two-thirds of all USDA tree pest control funds were devoted to efforts to suppress or eradicate the European gypsy moth (See FFI).
Concern about possible new introductions had grown; it focused on proposals to import unprocessed wood from Siberia, New Zealand, and Chile. The USDA Forest Service, academic scientists, and therefore APHIS emphasized the risks of known Asian pests, e.g., Asian gypsy moth, to western coniferous forests (See FFI). While individual scientists had expressed concern about wood packaging material, there was little public discussion of this threat.
We would learn later that several of the most damaging pests were already present in the country but not yet recognized – Asian longhorned beetle, sudden oak death pathogen, probably emerald ash borer.
beech leaf disease
photo by John Pogacnik
In 2019:
Numbers of non-native insects and pathogens attacking trees in North America approach 500 species. (In Fading Forests III, I calculated that by the first decade of the 21st Century, the number had risen to at least 475. Several more have been detected since 2014. More than 181 exotic insects that feed on woody plants had established in Canada. (Source: USDA APHIS. 2000. Wood packaging risk assessment.)
Of these, 91 are considered “serious” threats (Guo et al. 2019). This estimate excludes pests native to portions of North America that are causing severe damage in naïve hosts – e.g., goldspotted oak borer; pests of palms; and pests attacking trees on U.S. Pacific and Caribbean islands.
Introductions had continued.
Between 1980 and 2016, at least 30 non-native species of wood- or bark-boring insects (Scolytinae / Scolytidae) were newly detected in the U.S. (Haack and Rabaglia 2013; Rabaglia et al. 2019). A few of these are highly damaging, e.g. redbay ambrosia beetle, polyphagous and Kuroshio shothole borers.
In addition to these 30 new pests, other highly damaging tree-killing pests probably introduced since the 1980s include (on the continent):
Eight Cerambycids such as Asian longhorned beetle (Wu et al. 2017)
7 Agrilus, including emerald ash borer and soapberry borer; plus goldspotted oak borer transported from Arizona to California (Digirolomo et al. 2019; R. Haack, pers. comm.)
Sirex woodwasp
Pests of palm trees, e.g., red palm mite, red palm weevil, South American palm weevil
Spotted lanternfly
Beech leaf disease
Also not included in the above estimate and lists are tree-killing pests on America’s Pacific Islands :
‘ōhi‘a rust
Cycad scale
Cycad blue betterfly
Erythrina gall wasp
two Ceratocystis pathogens that cause rapid ‘ōhi‘a death
Coconut rhinoceros beetle
Authorities also carried out approximately 25 eradication programs targetting introductions of the Asian gypsy moth (USDA Pest Alert Asian Gypsy Moth plus additional outbreaks since 2014).
Impacts of exacerbated tree mortality rates linked to these introduced pests are seen across wide swaths of the country, and affect widespread species, genera, and families.
dead redbay in Claxton, Georgia photo by Scott Cameron
I will discuss the risk of continuing new introductions in a separate blog.
Trying to Develop
the Big Picture and Set Priorities
In
recent years, USDA Forest Service scientists have made several attempts to
provide nation-wide assessments of the impact of these pests and criteria for
establishing priorities.
The
National Insect and Disease Forest Risk Assessment predicted the loss of basal
area to various pests over the 15-year time period 2012 – 2027. The assessment
predicted the following losses for specific species: 90% for redbay; 60% for whitebark
pine; more than 40% for limber pine; 24% for tanoak; 11% for coast live oak; 6%
for eastern and Carolina hemlock; 27% for eight species of ash; 20% for
American elm; 19% for red oak; 18% for American beech (Krist et al. 2014).
A separate group of scientists found that, nation-wide, non-native forest pests are causing an approximate 5% increase in total mortality by tree volume (Randy Morin at NEFPC). For details on Dr. Morin’s findings, see my blog here.
A third approach to developing a nation-wide picture, Project CAPTURE, (and my blog here) utilized FIA data to develop priorities for conservation action. Fifteen species were placed in the highest priority category, including Florida torreya (Torreya taxifolia), American chestnut and Allegheny and Ozark chinquapins, redbay, five species of ash, two species of hemlock, Port-Orford cedar, tanoak, and butternut (Potter et al. 2019(b).
According
to Project CAPTURE, the non-native pests affecting the largest number of hosts
are the European gypsy moth, which attacks 65 hosts; and oak wilt (Bretziella fagacearum), which infects 61
hosts. The Asian longhorned beetle attacks 43 hosts (Potter et al. 2019(b).
I note that several other non-native pests also have high numbers of host species. In the Project CAPTURE study, these pests are ranked lower because the project limited its evaluation to the five agents with the greatest effect on any particular host. Thus, of the 18 native tree species that host one or both of the invasive shothole borers and associated Fusarium disease complex (PSHB website), the project included only six. Of the 22 tree species listed by APHIS as hosts of Phytophtora ramorum, the project included 12 (K. Potter, pers. comm. April 17, 2019).
SOD-killed tanoak on the Big Sur peninsula, California photo by Matteo Garbelotto, University of California Berkeley
More extensive discussions of
non-native pests’ impacts are provided in Lovett et al. 2006, Lovett et al.
2016, and Potter et al. 2019. A
book-length discussion of invasive species impacts – ranging from feral hogs to
invasive plants, is expected in December; look for Poland et al. (in press).
SOURCES
Aukema,
J.E., D.G. McCullough, B. Von Holle, A.M. Liebhold, K. Britton, & S.J.
Frankel. 2010. Historical Accumulation of Nonindigenous Forest Pests in the
Continental United States. Bioscience. December
2010 / Vol. 60 No. 11
Digirolomo, M.F., E. Jendek, V.V. Grebennikov, O. Nakladal. 2019. First North American
record of an unnamed West Palaearctic Agrilus (Coleoptera:
Buprestidae) infesting European beech (Fagus sylvatica) in New York
City, USA. European Journal of
Entomology. Eur. J.
Entomol. 116: 244-252, 2019
Guo,
Q., S. Fei, K.M. Potter, A.M. Liebhold, and J. Wenf. 2019. Tree diversity
regulates forest pest invasion. Proceedings of the National Academy of Sciences
of the United States of America. www.pnas.org/cgi/doi/10.1073/pnas.1821039116
Haack,
R.A. and R.J. Rabaglia. 2013. Exotic Bark and Ambrosia Beetles in the USA:
Potential and Current Invaders. CAB International 2013. Potential Invasive
Pests of Agricultural Crops (ed. J. Peña)
Krist,
F.J. Jr., J.R. Ellenwood, M.E. Woods, A. J. McMahan, J.P. Cowardin, D.E. Ryerson,
F.J. Sapio, M.O. Zweifler, S.A. Romero 2014. National Insect and Disease Forest
Risk Assessment. United States Department of Agriculture Forest Service Forest
Health Technology Enterprise Team FHTET-14-01
Leung,
B., M.R. Springborn, J.A. Turner, E.G. Brockerhoff. 2014. Pathway-level risk
analysis: the net present value of an invasive species policy in the US. The
Ecological Society of America. Frontiers of Ecology.org
Liebhold, A. M., W. L. MacDonald, D. Bergdahl, and V. C. Mastro. 1995. Invasion by exotic forest pests: a threat to forest ecosystems. Forest Sci., Monograph 30. 49 pp.
Lovett,
G.M., C.D. Canham, M.A. Arthur, K.C. Weathers, and R.D. Fitzhugh. Forest
Ecosystem Responses to Exotic Pests and Pathogens in Eastern North America. BioScience
Vol. 56 No. 5 (May 2006)
Lovett,
G.M., M. Weiss, A.M. Liebhold, T.P. Holmes,
B. Leung, K.F. Lambert, D.A. Orwig, F.T. Campbell, J. Rosenthal, D.G.
McCullough, R. Wildova, M.P. Ayres, C.D. Canham, D.R. Foster, SL. Ladeau, and
T. Weldy. 2016. NIS forest insects and pathogens in the US: Impacts and policy
options. Ecological Applications, 26(5), 2016, pp. 1437–1455
Mattson,
W. J., P. Niemela, I. Millers, and Y. Ingauazo.
1994. Immigrant phytophagous insects on woody plants in the United
States and Canada: an annotated list.
USDA For. Ser. Gen. Tech. Rep. NC-169, 27 pp.
Millers, I. United States
Department of Agriculture, Forest Service Entomologist, Forest Health
Protection Northeastern Area State and Private Forestry. Durham, NH. Personal
communication to F.T. Campbell, 1993.
Morin, R. presentation at Northeastern Forest Pest Council 81st Annual
Meeting, March 12 – 14, 2019, West
Chester, Pennsylvania
National Academy of Sciences. 1975. Forest Pest Control. Washington, D.C.
Poland,
T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019),
Invasive Species in Forests and Grasslands of the United States: A
Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. (in press).
Potter,
K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019. Important Insect and
Disease Threats to US Tree Species and Geographic Patterns of Their Potential
Impacts. Forests 2019, 10, 304.
Potter,
K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S. 2019. Prioritizing
the conservation needs of US tree spp: Evaluating vulnerability to forest insect
and disease threats, Global Ecology and Conservation (2019), doi:
https://doi.org/10.1016/
Rabaglia,
R.J., A.I. Cognato, E. R. Hoebeke, C.W. Johnson, J.R. LaBonte, M.E. Carter, and
J.J. Vlach. 2019. Early Detection and Rapid Response. A Ten-Year Summary of the
USDA Forest Service Program of Surveillance for Non-Native Bark and Ambrosia
Beetles. American Entomologist Volume 65, Number 1
U.S.
Department of Agriculture, Animal and Plant Health Inspection Service. 2009.
Risk analysis for the movement of wood packaging material (WPM) from
Canada into the US.
Wu,Y.,
N.F. Trepanowski, J.J. Molongoski, P.F. Reagel, S.W. Lingafelter, H. Nadel1,
S.W. Myers & A.M. Ray. 2017. Identification of wood-boring beetles
(Cerambycidae and Buprestidae) intercepted in trade-associated solid wood
packaging material using DNA barcoding and morphology Scientific Reports 7:40316
In January I wrote about possible changes to the 2018 Farm Bill aimed at restoring tree species devastated by non-native insects and diseases. (Earlier, in July 2017, I had blogged about these and other proposed amendments to the Farm Bill. Here’s an update.
The House Committee on Agriculture has completed action on its version of the Farm Bill; it now awaits action by the full House. The House bill (H.R. 2) does not contain any of the proposals put forward by the Center for Invasive Species Prevention to improve both “rapid response” to introduced forest pests or long-term efforts to restore species devastated by such pests. It does contain very controversial provisions weakening the Endangered Species Act, National Environmental Protection Act, and Federal Insecticide, Fungicide and Rodenticide Act. The bill also makes major changes to the food stamp program. Most political analysists do not think the bill will become law in its current form.
We do not yet know what provisions will be in the Senate’s version of the Farm Bill. I am still hopeful that our proposals might be included in some form.
Rep. Peter Welch
Meanwhile, Representative Peter Welch from Vermont has introduced a separate House bill: H.R. 5519. It would amend the Plant Protection Act in order to increase funding for both APHIS-led programs to eradicate or contain invasive species and for competitive grants to research specific questions related to the recovery of pest-decimated native tree species.
Specifically, Mr. Welch’s Invasive Species Prevention and Forest Restoration Act would:
Strengthen the USDA Secretary’s authority to access Commodity Credit Corporation funds to supplement appropriations to counter non-native insects and pathogens and noxious weeds that threaten agricultural production (including forest resources).
Create a competitive grant program to address the critical threat to numerous tree species posed by non-native plant pests and noxious weeds. The grants would provide up to $100,000 per year, over two to five years, to support research that promotes the restoration of affected tree species, including research on—
(A) biological control of plant pests or noxious weeds;
(B) exploration of genetic manipulation of plant pests or noxious weeds;
(C) enhancement of host-resistance mechanisms; and
(D) development of other strategies for restoring tree species.
These studies must be part of comprehensive forest restoration research.
The research institutions are to disseminate to the public tools and information based on the research conducted under this program.
Institutions eligible for funding under this program include:
(1) An agency of the Federal Government.
(2) A State cooperative institution.
(3) A college or university offering a baccalaureate or higher degree in the study of food, forestry, and agricultural sciences.
(4) A nonprofit entity described in § 501(c)(3) of IRS Code.
A committee of government experts would advise the Secretary of Agriculture on applying the funding criteria for the grant program. This committee would be composed of representatives of the USDA Forest Service, APHIS, and Agriculture Research Service; and State forestry agencies. This committee of experts would receive advice from a separate advisory committee composed of representatives of land-grant colleges and universities, affiliated State agriculture experiment stations, the forest products industry, recreationists, and conservation organizations.
Funds from the Commodity Credit Corporation (not subject to annual appropriations) are made available, beginning at $3 million in Fiscal Year 2019 and rising to $10 million in FY 2022.
Please ask your Representative to co-sponsor this bill. Also, ask your Senators to consider sponsoring a similar bill in the Senate (assuming that no such provisions are included in the Senate Farm Bill).
If these proposals are not included in the version of the Farm Bill that is adopted this year, there might be new opportunities to advance resistance breeding and similar programs in the future. As I noted in the March blog, a growing number of scientists and concerned stakeholders have developed proposals to expand resistance breeding programs. I hope that the several groups can coordinate their efforts and recruit supporters for a lobbying effort that will create a coordinated program to restore native trees devastated by non-native insects and diseases.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.



















 Port-Orford cedar resistance trials
Port-Orford cedar resistance trials