Science continues to provide new hope for management of some of the tree-killing insects introduced to North America.
searching for ALB; photo by USDA
Asian longhorned beetle
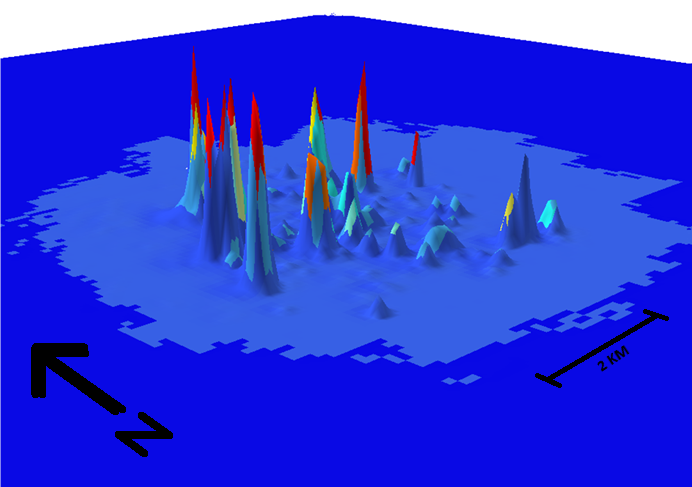
USDA Forest Service and APHIS scientists have developed an Asian Longhorned Beetle (ALB; Anoplophora glabripennis) Eradication and Risk Tracking (ALBERT) 1.0 toolkit. Trotter et al. (2024) believe that the toolkit will improve the efficiency of ALB eradication programs by better estimating changing infestation risk in real time and comparing efficacy of various management strategies in those circumstances. Managers can thus set priorities for surveys and eradication efforts, optimize surveys, and improve eradication outcomes. Improved efficiency will also reduce long-term management costs.
Data on dispersal and infestation risk are analyzed at the hectare level. Inputs are contemporary, so they reflect hectare-scale changes in risk level and ALB detectability simultaneously with program implementation. Managers and stakeholders can track and visualize the distribution of risk on the landscape. In the example, the risk of infestation is shown by the color and height of the surface on the landscape.
I note that the research was completed two years ago. I wish the USFS “Rooted in Research” brief had discussed whether programs managing extant ALB infestations are using the tool – and finding it useful.
Suppressing emerald ash borer populations
black ash swamp via Flickr
Minnesota authorities and scientists wish to protect the expansive black ash (Fraxinus nigra) swamps located mostly in the central & northern portions of the state. There, the emerald ash borer (EAB; Agrilus planipennis) has not yet spread to all vulnerable locations. They consider that use of pesticides is not concerned feasible in these locations. And two of the introduced biocontrol agents have limited efficacy. One of the wasps—Tetrastichus planipennisi— can oviposit in EAB larvae only in trees with thin bark, that is trees smaller than 16 cm dbh. A second wasp, Spathius agrili, doesn’t persist in northern regions – i.e., northern Minnesota! Therefore they are seeking additional management tactics workable in natural mature ash stands across northern North America.
Peters, Rajtar, & Blanchette (2025) are testing whether useful levels of suppression can be provided by release of entomopathogenic fungi (EPF). They are building on earlier studies link? that determined that one entomopathogenic fungus isolate, Beauveria bassiana, showed promise in reducing EAB population growth. This fungus is available as a commercial product.
Peters, Rajtar, & Blanchette (2025) tested isolates from five genera = Beauveria, Purpureocillium, Metarhizium, Clonostachys, & Samsoniella. They found that Beauveria pseudobassiana, Beauveria bassiana GHA, Metarhizium sp. Meta, & Purpureocillium sp. consistently reduced EAB mean survival time (MST) and probability of survival over time.
They also tested use of an inoculation method different from that used in the earlier studies: autodissemination devices (ADDs). The devices expose adult EAB to the fungal spores using baited Lindgren funnels with non-stick surfaces hung in the canopy of an infested ash tree. Infected beetles transmit the fungus to others during mating. In a Canadian study, this method of infecting EAB adults with an isolate of B. bassiana CFL-A caused rates of infection up to 40% and a significant decrease in EAB pop growth rate over a 3-year period.
Scientists have isolated and identified 1,126 fungal isolates associated with EAB larval galleries from trees growing across Minnesota. Eight percent of these isolates are entomopathogens. Some of these isolates have been proved to kill EAB eggs. In this study Peters, Rajtar, & Blanchette (2025) tested the fungi’s efficacy against adults and whether the ADDs system inoculates sufficient numbers of beetles.
They found that five of the fungal isolates significantly decreased the mean survival time of EAB adults post-exposure and the probability of EAB survival over time. These were Beauveria bassiana GHA, B. pseudobassiana EAB 16.8 & B. pseudobassiana EAB 53-5, Metarhizium sp. Meta, and Purpureocillium sp. EAB 59-16-2. They are most promising isolates for future study. Peters, Rajtar, & Blanchette (2025) discourage further consideration of isolates in the genera Samsoniella and Clonostachys because of disappointing reductions in adult beetle survival.
The probability of survival of beetles in the negative control groups differed significantly between the two experiments. The authors say they can only speculate about the reason because many external factors influence EAB adult lifespan. Among such factors are a) the longer storage period for insects used in the first experiment; b) differences in nutrient quality or microbiomes of ash leaves provided during incubation; or c) health of trees from which the beetles had been collected.
Peters, Rajtar, & Blanchette (2025) say the effectiveness of these fungi and this inoculation method must now be tested under field conditions. One concern: the fungal inoculum can degrade due to exposure to the environment while it is in the trap. A second concern is that the fungus might grow across the inoculum pouch unevenly, thereby undermining equal delivery of conidia to each beetle. They call for research to optimize the type and/or formulation of inoculum used in ADDs to prolong fungal viability in the field.
an ash in a suburban setting; photo by F.T. Campbell
While I am cheered by the prospect of conservationists having new tools to counter the EAB infestation, I am concerned by how slowly work pursuing possible use of entomopathogens is proceeding. Peters, Rajtar, & Blanchette (2025) refer to several past studies that reached similar findings. Why has this knowledge not been applied in the field already? Is it due to the intrinsic difficulty of detecting and working with the fungi? Lack of funding for non-commercial approaches? Leaders’ narrow focus on strategies already in use? Some other complication?
SOURCES
Peters, C.J.; Rajtar, N.N.; Blanchette, R.A. Entomopathogenic Fungi from MN Are Virulent Against Emerald Ash Borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae), Adults in a Laboratory Autodissemination Device Assay. Forests 2025, 16, 1742. https:// doi.org/10.3390/f16111742
Trotter, R.T., III; Warden, M.L.; Vazquez, R.J.; Ryan, J.K.; Pfister, S. 2024. ALB Hazard Management and Monitoring Version 1.0: an assessment and tracking tool for Asian longhorned beetle eradications in the United States. Gen. Tech. Rep. NRS-222. Madison, WI: U.S. Department of Agriculture, Forest Service, Northern Research Station. 24 p. https://doi.org/10.2737/NRS-GTR-222.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Barbours Cut terminal, Port of Houston; photo by Patrick Feller via Flickr
I blogged recently about ports in the southeast dredging channels and taking other actions to increase the number of container ships they can service. Michael Angell of the Journal of Commerce reports that the Port of Houston is competing vigorously for visits by ships from Asia. Of course, rising ship visits increase the opportunity for Asian pests such as Asian longhorned beetle or several ambrosia beetles in the Euwallaceae complex to invade the Gulf Coast region. (The emerald ash borer is already present in Texas.)
Considering the immediate future, the Port of Houston is expanding capacity of its two existing terminals. The Port is dredging portions of its channel down to a 46-foot depth. It expects to complete dredging of the main channel to 55 feet – the depth of Norfolk currently by ten years from now.
One of the existing terminals, the Bayport container terminal, handles most of the trans-Pacific Asia services coming into the Gulf Coast. Two new berths and storage yard space are being added, so that Bayport will be able to handle five super-post-Panamax ships simultaneously. The other terminal, Barbours Cut terminal, is also being renovated to improve handling of super-post-Panamax ships. At full buildout, the Bayport and Barbours Cut will have capacity for handling 8 million TEUs per year.
Houston has begun design work for a third container terminal. At full capacity – more than a decade from now — this third terminal will handle another 4 million TEUs. The project has been approved by the Federal Permitting Improvement Steering Council for streamlined environmental under the 2015 FAST Act. This program does not alter any applicable statutory or regulatory requirement, or guarantee approval. It seeks to expedite lawfully mandated environmental and other review through standardized interagency consultation and coordination practices. It also created a public online tool to track progress in completing these requirements.
Through May 2026, Houston handled 502,387 TEUs in container imports from Asia, up 10% from the same period in 2025. The number of container ships over 10,000 TEUs in capacity calling at Houston doubled, from four in the second half of 2025 to nine during the first half of 2026.
Port of Long Beach
Houston ranked fifth in container volumes during this period. Los Angeles was the top port, with 1,974,330, followed closely by Long Beach with 1,851,159. Together, the two California ports received more than seven times as many containers as Houston. Houston is competing with other ports in the Southeast. Savannah still handles more than a third higher numbers than Houston, at 823,823 containers. The Port of Virginia (see earlier blog) received 285,046 containers, and Charleston only 256,499 Both were decreases from the 2025 number. All three southeastern ports saw small decreases from their 2025 numbers.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Tillandsia utriculata; photo courtesy of Teresa Cooper
Florida has 16 native species of bromeliads. Up to 15 invertebrates depend on bromeliads, especially the water that collects at the bases of the leaves. See Frank (1983) and Frank and Fish (2008) for more information about bromeliads’ ecological importance.
Eight species had been listed as threatened or endangered in the state (visit here) (Frank and Cave 2005) before arrival of the Mexican bromeliad weevil. The Florida Endangered Plant Advisory Council added two of its hosts – Tillandsia utriculata and Tillandsia fasciculata. The website now lists 12 species as threatened by the weevil.
Twelve species are believed to be vulnerable to the weevil. T. utriculata is at most immediate risk. Before arrival of the Mexican bromeliad weevil, some populations of this species were described by Teresa Cooper as “giant” and dense, containing very large mature plants and producing huge volumes of of seed.
One such population — in the Enchanted Forest Sanctuary in Brevard County – succumbed to the weevil quickly: 87% died in six months. At 27 months after arrival of the weevil, less than 3% of the original population remained. It is now rare to see large T. utriculata plants. Medium-sized plants put out inflorescences. The weevil persists at low levels, so is able to feed on and reproduce on medium and small plants. Several endemic wildlife species that inhabit the water pooled in T. utriculata’s base face global extinction (T.M. Cooper pers. comm. 2026).
damage to a bromeliad caused by the weevil; photo courtesy of Teresa Cooper
The first reason T. utriculata is at higher risk is that each plant can support many weevil larvae. Its leaves are softer (so more easily consumed) and its large stems contain lots of nutrients — for a bromeliad). More important is the plant’s very slow reproduction in contrast to the weevil’s rapid attack. T. utriculata plants do not produce seed until after they have grown for up to 20 years. Furthermore, an extremely small fraction of the ~ 10,000 seeds released by a mature plant make it to reproductive age. A plant that after 18 years begins growing an inflorescence — which then takes ~ a year to flower and produce seed – can be killed by a weevil within a few months. In addition, T. utriculata is monocarpic – it doesn’t produce offshoots. T.M. Cooper asserts that if the bromeliad weevil extirpates T. utriculata from Florida, it will not be because the weevil ate the very last plant, it will be because T. utriculata seed output has fallen below a sustainable level.
Teresa Cooper demonstrating the size of a T. utricuclata flower spike; photo courtesy of T. Cooper
A second host, Tillandsia fasciculata, had large and dense population in Loxahatchee National Wildlife Refuge (Palm Beach County) in 2002 to 2005. Almost every tree had multiple T. fasciculata plants; many were huge with multiple pups. When Teresa Cooper revisited about 10 years later, T. fasciculata plants were very sparse, large plants very rare; none was really large (T.M. Cooper pers. comm. 2026).
T. fasciculata is expected to decline more slowly than T. utriculata. First among several advantages is that its leaves are tougher and stems smaller, so they provide less nutrition to the weevil. Probably more important, T. fasciculata is polycarpic (it produces both seed and offshoots). A large T. fasciculata plant can sprout up to a dozen blooming rametes at a time. If the weevil kills one, other rametes will bloom and seed. At the same time, the plant might produce numerous up to 20 offshoots which come detatched from the “mother” plant and start a new individual. Still, as the weevil kills these rametes year after year the plant shrinks in size and eventually dies (T.M. Cooper pers. comm. 2026).
Teresa Cooper has documented damage on Tillandsia paucifolia, T. simulate, and T. variabilis (T.M. Cooper pers. comm. 2026). She has no funding to continue studying these species. I have been unable to find any more recent information about the status of the Tillandsia species. The Florida Department of Agriculture and Consumer Services does not have an active program addressing the weevil (Patrick Gordon, FDACS, pers. comm. June 2026).
There is considerably more information about another endangered bromeliad, Guzmania monostachia. This species has the broadest distribution of any species in the genus, stretching across northern South America, Central America, the Caribbean, as well as southern Florida. As the northernmost population, the Florida population might harbor unique genetics important for the species’ adaptation to climate change (Krupar et al. 2023).
Formerly more common, G. monostachia is now found in six fragmented and disjunct populations in five areas in Florida. The species has apparently been extirpated from four other sites by habitat loss, not depredations by the weevil. One set of habitats is in wetland sloughs on the peninsula’s west coast on the peninsula’s west coast. These forests comprise pond apple (Annona glabra) and Florida ash (Fraxinus caroliniana); Kupar et al. (2023) don’t mention whether this species is vulnerable to the emerald ash borer, which has not yet been detected in Florida. The largest of the bromeliad populations is in Fakahatchee Strand State Preserve, in Collier County in Southwest Florida. Fakahatchee Strand Preserve is Florida’ oldest and largest state park. The bromeliad’s population was previously estimated to exceed two million individuals. Their number has been halved by weevil attack. Also, since M. callizona prefers larger plants, the proportion of reproductively mature individuals had been reduced from roughly 50% to only 10–20% by 2021. T.M. Cooper reports (pers. comm. 2026) fears this species might also be extirpated eventually.
Guzmania monostachia; photo courtesy of Teresa Cooper
The adjacent Big Cypress National Preserve is home to two populations of G. monostachia; they are separated by ~2,900 km2 so there is no interaction between them. The southern population consists of ~ 1000 individuals, the northern population comprises only~ 200 individuals.
Guzmania monostachia is in an even more precarious situation on the eastern side of the peninsula: tiny populations of two or three plants are found in three locations: Everglades National Park, Fuchs (formerly Sykes) Hammock Preserve, and Meissner Hammock Preserve.
The Mexican bromeliad weevil Metamasius callizona (Chevrolat) is native to southern Mexico, Guatamala, (Frank and Thomas 1994; Frank and Cave 2005) and Belize (Cooper, Cave and Frank 2023). Its presence in Florida was detected in a nursery in Ft. Lauderdale, Broward County, in 1989. Probably introduced on a shipment of ornamental bromeliads from Mexico. The nursery treated the infested plants, but the weevil had already established on native bromeliad species in the natural environment. By 1991, it was detected in four counties in southern Florida; by 1999, it was found in 12 more. It is now in most counties of peninsular Florida from Daytona (Volusa County) south to Miami-Dade (Patrick Gordon, Florida Department of Agriculture and Consumer Services, pers. comm. June 2026).
While chemical control is feasible in nurseries and display plantings, it can’t be used in natural areas, where the epiphytes are not accessible from the ground and stakeholders fear likely non-target effects.
Therefore, scientists initially focused on classical biocontrol. They made 16 expeditions to Mexico and Central and South America from 1992 to 2010 searching for natural enemies of the weevil. The chose parasitoid tachinid fly, Lixadmontia franki. A colony was established in the University of Florida’s quarantine laboratory and research on fly-weevil interactions was conducted for several years. Releases into the environment were begun under Federal and State permits in 2007. Although more than 3,100 flies were released, the species failed to establish. So this approach is no longer being pursued (Cooper, Cave, and Frank (2024); T.M. Cooper pers. comm. June 2026).
Meanwhile, in 2010, Dr. Frank and D. Giardina of the Florida Fish and Wildlife Conservation Commission observed that in one location in Belize the weevils had no apparent detrimental effect on bromeliads identified as Tillandsia utriculata (although there is some disagreement on this classification). Cooper, Cave, and Frank (2024) undertook a study to determine how the bromeliad could coexist with Metamasius callizona in Belize while being so vulnerable in Florida. They compared life cycle parameters (oviposition and pupation rates, egg hatch rate, adult emergence and size, and developmental time) of weevils from Florida and Belize. They also observed how populations of the weevil from Florida fared on three hosts: pineapple (Ananas comosus), T. utriculata collected from Florida, and T. utriculata collected from Belize. Finally, they quantified the hosts’ nutritional content using two measures (percent soluble solids and leaf toughness).
Their most important finding is that weevil larva from Florida could not develop past the third instar on leaves of the T. utriculata from Belize. The weevil larvae starved because the Belize bromeliad’s leaves had fewer nutrients and were tougher than the leaves from T. utriculata collected in Florida.
bromeliads seen in Big Cypress National Preserve in the early 1980s; photo by F.T. Campbell
Based on this finding, Cooper, Cave, and Frank (2024) recommend that authorities introduce T. utriculata plants from Belize into Florida’s forests and allow them to colonize and/or hybridize naturally with Florida’s remaining plants. An alternative would be to breed hybrid Tillandsia in the lab and release them into Florida’s forests. I note that the American Chestnut Foundation pursued a similar strategy for decades to develop American chestnuts (Castanea dentata) able to tolerate the chestnut blight fungus (Cryphonectria parasitica).
Sources
Cave, R.D. 1997. Admontia sp., a potential biological control agent of Metamasius callizona. J Brom Soc. 47:244-249.
Cave, R.D. 2008. Biological control of the MEXICO bromeliad weevil. Biocontrol News and Information 29(1):1N-2N.
Cooper T.M. 2006. Ecological and demographic trends and patterns of Metamasius callizona (Chevrolat), an invasive bromeliad-eating weevil, and FLORIDA’s native bromeliads [Master’s thesis]. [Gainesville (FLORIDA)]: University of FLORIDA. 69 p.
Cooper, T.M., R.D. Cave, and J.H. Frank. 2023. Potential bottom-up control of Metamasius callizona in Florida, USA. Entomologia Experimentales et Applicata. 2024. 172;4090421
Frank, JH. 1983. Bromeliad phytotelmata and their biota, especially mosquitoes. In: Frank J.H., Lounibos, L.P., editors. Phytotelmata: terrestrial plants as hosts for aquatic insect communities. Medford (NJ): Plexus. p. 101-128.
Frank, J.H. and R.D. Cave. 2005. Metamasius callizona is destroying FLORIDA’s native bromeliads. In: Hoddle MS, editor. USDA Forest Service Publication FHTET-2005-08. Vol 1. Second International Symposium on Biological
Control of Arthropods; 2005 Sep 12-16; Davos, Switzerland. Washington D.C.: USDA Forest Service. p. 91-101.
Frank, J.H. and D. Fish. 2008. Potential biodiversity loss in Florida bromeliad phytotelmata due to Metamasius callizona (Coleoptera: Dryophthoridae), an invasive species. Florida Entomol. 91(1):1-8.
Frank, J.H. and M.C. Thomas. 1994. Metamasius callizona (Chevrolat) (Coleoptera:Curculionidae), an immigrant pest, destroys bromeliads in FLORIDA. Can Entomol.126(1):673-682.
Krupar, S., A.A. Naranjo, G. Godden, N. Cellinese. The Fate of Guzmania monostachia in Florida Rests with Humans. Diversity 2023, 15, 525. https://doi.org/10.3390/d15040525
Potter, K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S., Prioritizing the conservation needs of US tree species: Evaluating vulnerability to forest insect and disease threats, Global Ecology and Conservation (2019), doi: https://doi.org/10.1016/
Salas, J. and J.H. Frank. 2001. Development of Metamasius callizona (Coleoptera:Curculionidae) on pineapple stems. Florida Entomol. 84(1):123-126.
Wood, D.M. and R.D. Cave. 2006. Description of a new genus and species of weevil parasitoid from Honduras (Diptera: Tachinidae). Florida Entomol. 89(2):239-24.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Cibotium glaucum in Hawaii Volcanoes National Park; F.T. Campbell
A year ago I blogged about the status of Hawaiian species in the face of high levels of bioinvasions. I now update that discussion as it pertains to one of the invaders, the Australian tree fern Sphaeropteris cooperi.
S. cooperi has successfully invaded multiple regions, including the southern coast of South Africa, the Azores and Mascarene Archipelagos, and Hawai`i. The invasion’s impacts have been assessed only in Hawai`i. Studies found that the non-native tree ferns have outcompeted the dominant native tree fern Cibotium glaucum (Cibotiaceae), and altered leaf litter composition hence soil nutrient cycling. This in turn affects species assemblages (van den Berg et al. 2025).
Chau, Walker and Mehltreter (2013) documented the litter and soil chemistry in Hawaiian rainforests. They found that S. cooperi produces more leaves that grow faster, contain more nitrogen and phosphsorus, and decompose faster than the leaves of the dominant native tree fern C. glaucum. They compared the effect of leaf litter from the native and tree ferns on the growth and nutrient content of four native angiosperm species when they were grown in N-rich forest soil and P-rich landslide soil. The results suggested that nitrogen availability is the strongest driver of growth. The introduced tree fern, S. cooperi, can thus prompt more rapid growth of some native HI plants. They point out, however, that under natural conditions, native plants must compete for these additional nutrients with various non-native plants, including S. cooperi. The ultimate impact, then, remains unclear.
van den Berg et al. (2025) report that other invaded sites are at great risk Both the Garden Route region of South Africa and La Reunion Island in the Mascarene Archipelago in the eastern Indian Ocean have high bioidiversity. Both contain large, globally-recognized protected areas established to protect the native biodiversity: UNESCO Garden Route Biosphere Reserve and Reunion National Park. Despite their biological importance, both are among the top10 most invaded countries/territories globally (Tuberlin et al. 2017).
Sphaeropteris cooperi; photo via Easyscape
While climate change is expected to reduce the extent of suitable habitat for the Australian tree fern in both South Africa and La Reunion, the current situation is troubling. At present the tree fern occupies a narrower range of climatic conditions in both the Garden Route and especially on La Reunion than in Australia. (On La Reunion, 13.74 % of the fern’s apparent niche remains unoccupied.) van den Berg et al. (2025) are not sure what factors might be limiting the tree fern’s spread. They do urge educational campaigns to persuade people living near the Biodiversity Reserve to avoid planting the non-native tree fern.
The tree fern’s actual niche is somewhat uncertain because predation by deer (non-native to Australia) might have reduced its reproduction. The authors mention this but do not speculate further on the possible response of the fern to the absence of such stresses on La Reunion.
SOURCES
Chau, M.M., Walker, L.R. and Mehltreter, K. An invasive tree fern alters soil and plant nutrient dynamics in Hawaii. Biol Invasions15, 355–370 (2013). https://doi.org/10.1007/s10530-012-0291-0
Turbelin,A.J., Malamud,B.D., Francis,R.A. 2017. Mapping the global state of invasive alien species: patterns of invasion and policy responses. Glob. Ecol. Biogeogr. 26,78–92. https://doi.org/10.1111/geb.12517.
van den Berg, M.L., G. Singh, E.J. McCulloch-Jones, M. Rouget, D.M. Richardson, T.B. Robinson. 2025. The invaded range of the tree fern Sphaeropteris cooperi is predicted to shrink in two southern hemisphere biodiversity hotspots. African Journal of Botany 178 (2025) 390-399
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
dying ash in Shenandoah National Park; photo by F.T. Campbell
The emerald ash borer (Agrilus plannipennis; EAB) was detected in North America in 2002. So both U.S. and Canadians have been motivated to evaluate the probable trajectory of the primary hosts – the ash genus Fraxinus – in the face of the ongoing invasion. See Deschênes et al. 2026 and Wilson et al. 2025 – full citations at the end of this blog. Both studies focused on white (Fraxinus americana) and green ash (F. pennsylvanica); they say next to nothing about black ash (F. nigra). I regret this silence because of the unique ecology of black ash swamps. Neither addresses the threat to Oregon ash (F. latifolia) in the West.
The two assessments have similar findings: high mortality of larger trees (canopy and “recruit” size trees); abundant regeneration (seedling and saplings sizes) after an initial period; and uncertainty as to whether persisting EAB populations will kill the saplings before sufficient numbers grow into reproductive size.
There are two conspicuous differences. First, the American study does not consider the possible impact of biological control – although USDA APHIS has placed all its effort on this approach since January 2021. The Canadians report that self-sustaining populations of the wasps are now found across the northern U.S. and eastern Canada. In their study, conducted in Ontario, they detected only Tetrasticus planipennisi; it was parasitizing 16% of the EAB larvae in dissected stems. This wasp’s affinity for colder climates and short ovipositor – which limits it to parasitizing larvae inhabiting small stems – are portrayed as positive traits under these circumstances.
Second, the Canadians did not find “lingering” adult ash trees as have the Americans. These trees indicate the probability of finding workable levels of genetic resistance to the EAB. USDA Forest Service scientists are pursuing a breeding program. While in south-central Michigan, where overstory ash mortality typically exceeded 80%, 46% of overstory ash and 82% of ash recruits were relatively healthy (Wilson et al. 2025), in Ontario none of 1,129 overstory ash trees survived beyond seven years after EAB detected. No trees exceeded 15 cm dbh (Deschênes et al. 2026).
ash resistance breeding trial at Holden Arboretum; photo courtesy of Jennifer Koch, USFS
Regeneration
As Deschênes et al. (2026) state, the future of ash stands depends on the complex of interactions among environmental conditions, management interventions, efficacy of natural enemies (natural or introduced), and life-history traits of the insect and its host. Coexistence might be possible if EAB larval densities remain sufficiently low to support survival of residual trees and successful seedling recruitment.
Larval densities in Ontario were said to be generally low, suggesting reduced carrying capacity in post-invasion forests, lower EAB fecundity, and higher EAB mortality in regenerating stems. Deschênes et al. (2026) note that in more northern areas colder temperatures are thought to slow larvae development. Perhaps these larvae might also be less vigorous, so they night disperse only over short distances. Still, there were sufficient EAB present after all the overstory trees had died to create 97% of the 298 galleries in regenerating ash stems (Deschênes et al. 2026). Furthermore, Wilson et al. (2025) say that EAB densities in infested trees in Michigan were similar to densities recorded during the initial invasion. This seems ominous to me – a solid foundation from which beetle populations could build up again as regenerating ash grow and provide more phloem for the insect to exploit.
Ash reproduce by both flowering/seeding and sprouting from the base. EAB predation is not the only complication. First, ash are dioecious so mature trees of both sexes must grow within a few hundred meters. Second, predation by the ash seed weevil (Lignyoodes helvolus) reduces seed supplies. Dense sedge mats can prevent germination (Wilson et al. 2025). Scientists generally believe that the soil seed bank is quickly exhausted, although Wilson et al. (2025) cite others’ conflicting findings. Neither article discusses predation by mammals, e.g., deer or rabbits. Wilson et al. (2025) mention attacks by beavers.
ash saplings felled by beavers; photo by F.T. Campbell
Wilson et al. (2025) did not study whether stump-sprouted ash were able to successfully recruit into the overstory. They do report that in one study in southeastern Michigan stump sprouts were the dominant form of green ash regeneration and about a quarter of these sprouts produced seeds at least once. Deschênes et al. (2026) found that on average 47% of regenerating stems at their Ontario research sites originated from stump sprouts.
EAB has been documented to attack and kill trees when the main stem is as small as 2.5 cm. While EAB probably prefer larger stems, Deschênes et al. (2026) suggest that stems become acceptable at the lower range of size required for seed production – 8–10 cm dbh. Reliable and abundant seed production doesn’t occur until white or green ash achieve > 20 cm dbh. At their Ontario sites, Deschênes et al. (2026) found that 42% of regenerating stems has been infested by EAB at least once; 14% had been attacked five or more times. They removed 74 EAB larvae from 28 stems; 49 (66%) were alive. Fifteen EAB (16% of current year galleries) had been parasitized — all by Tetrastichus planipennisi. They also observed numerous signs of defensive responses.
In Michigan, no ash recruits — living or dead – were found in plots in 28% of the cells. In the remaining 128 cells, an average of ~33% of ash recruits were infested by EAB, and ~21.4% of ash recruits dead. As is typical, white ash fared better than green ash. Recruit sized ash trees were twice as likely to die than to survive and mature into overstory size (Wilson et al. (2025).
In Ontario, as noted, all canopy ash had died. There were 119 live trees 5 – 10 cm dbh – a tenth as many “mature” ash as when EAB arrived, and all were smaller. There was abundant regeneration in most sites initially, but at the longest-infested sites in Essex County, regenerating ash stems were half as numerous as early after the transition (Deschênes et al. 2026).
The Canadians found it encouraging that some of the regenerating stems were vigorous despite containing EAB gallery densities greater than 20 larvae·m?2 of phloem. They did not know the mechanisms underlying survival of these stems. Possible explanations ranged from the low EAB carrying capacity of smaller trees to stronger host defenses in regenerating stems to EAB mortality due to parasitism.
Wilson et al. (2025) note that despite more than 20 years of EAB presence, densities of ash recruits, saplings, and seedlings were high relative to other species. However, they remind us, ~ one-third of the live ash recruits were infested so their survival into reproductive size was uncertain. The high mortality of overstory ash results in loss of seed resources, greater sun exposure, and cascading consequences for forest composition and function. In upland sites, cells with low ash basal area favored Quercus rubra and Tilia americana. They conclude that changes to forest composition is probably site specific — largely depend on what tree species are already present.
Despite the challenges described above, the Canadian scientists also believe that these findings demonstrate that ash has a capacity for long-term regeneration (Deschênes et al. 2026).
Changing Species Composition in the U.S. (Wilson et al. 2025)
Canopy gaps caused by ash mortality have largely been filled by lateral ingrowth of species already there — American elm (Ulmus americana), black cherry (Prunus serotina), and northern red oak (Quercus rubra). The regeneration strata (saplings and seedlings) is dominated by Fraxinus (white outnumbering green when differentiated), maples (Acer rubrum, A. saccharum), black cherry, Crataegus species and Carya ovata. Elms are consistently among most common non-ash taxa among overstory, recruit, sapling and seedling strata. At some Ohio sites there was also increased abundance of non-native tree and shrub seedlings. This is not surprising since invasive plants are widespread in the forests of Ohio and other eastern states. A decade ago 93% of Forest Inventory and Analysis (FIA) plots in Ohio had at least one of 50 invasive plant species.
In another paragraph they mention Tilia americana as one of the important species in these forests.
Situation in Canada (Deschênes et al. 2026)
Deschênes et al. (2026) express concern that the death of nearly all canopy-level trees will substantially reduce ash’ ability to fulfill its ecological roles in these ecosystems. Still, ash regeneration is persisting for decades following overstory mortality. The taxon’s continued presence is driven largely by strong sprouting, which has been observed in several locations in Ontario. In some areas, low EAB infestation rates and evidence that regenerating stems can withstand multiple infestations raises hope that some might reach maturity and produce seeds. This scenario would be similar to that of elms, in which surviving trees contribute to ongoing regeneration and might eventually facilitate development of some level of resistance to the invasive fungus. A second possibility is that ash’ high sprouting capacity might point to a scenario similar to that of American chestnut. This species has persisted for a century primarily as sprouting shrubs — although they rarely reach reproductive maturity.
white fringetree; photo by Ryan Somma via Wikimedia
White Fringetree
Scientists also reviewed the status of a secondary host of EAB in North America, white fringetree (Chionanthus virginicus). Earlier studies of this host-pest relationship had been conducted on ornamental plantings where the trees tend to be scattered across open lawns and actively managed – including protection from pests. The Cipollinis (see full citation at the end of this blog) believe they might be better able to ward of EAB attack than are wild, unmanaged trees in forests that must compete for resources. They wanted to assess the current status and likely trajectory of the tree species in the wild.
To do so they revisited a wild population of the tree in southern Ohio previously assessed 10 years earlier. White fringetree is a small multi-stemmed tree native to the southeastern U.S. It is widely planted as an ornamental in across the east. In Ohio, white fringetree grows wild in only a few southern counties, in small populations or as widely scattered individuals. The species is classified as “Potentially Threatened” at the state level.
In 2015, 30% of the white fringetrees at the site were infested. These trees had signs of stress but none had died. EAB larvae grow more slowly on fringetree than on North American ash species. Meanwhile, all mature white ash trees at the site had been killed by EAB. Smaller white ash trees more comparable in size to the white fringetrees had attack rates and impacts comparable to those on the fringetrees.
In their new study, the Cipollinis found that nine of 31 trees tagged in 2015 (29%) had died; 22 (71%) were alive. Of those 22 living trees, 12 (55%) stayed the same or improved slightly over the five-year period; 10 (45%) declined. Five of these 22 living trees (23%) had evidence of current infestation. Trees that had died had a higher incidence of old EAB galleries, adult exit holes, and woodpecker activity. This is interpreted as demonstrating that EAB must cause extensive damage to kill fringetrees.
In summary, fringetrees in a wild unmanaged population continued to be attacked by EAB over 10 years and suffered higher attack rates and more significant impacts than those previously observed in managed pops. The Cipollinis conclude that trees large enough to attract EAB oviposition will continue to decline in health and be killed as long as beetles are present. They expect that wild white fringetrees might meet the same fate as ash trees, but over longer time scales.
At the same time, this delay in complete mortality might create a refugium for remnant populations of EAB after most ash have been killed. This status would be exacerbated if it turns out that the biocontrol agents cannot find their target — EAB — in the alternative host. The Cipollinis found lower parasitism rates by Tetrastichus planipennisi in fringetrees, although this was not true for the egg parasite Oobius agrili and two Spathius larval parasites.
Whitebark fringetree populations can produce few adult EAB because the trees are small and contain low amounts of phloem. Still, as young trees grow into vulnerable sizes they might help sustain the EAB population – as young ash trees in the area appear to do.
While caution is appropriate in interpreting findings from a study of a single population, the Cipollinis argue that this population has been studied intensively: assessed six times over 10 years, beginning at the start of the EAB infestation. Therefore they think their analysis provides useful informative regarding the long-term impacts of EAB on fringetree.
They concede that larger populations in areas deep within the tree’s native range might experience different dynamics and impacts. So far, however, observations in Chattahoochee National Forest in Georgia and at Great Falls Park on the Maryland-Virginia border generally support their finding that wild fringetrees in natural landscapes will suffer higher attack rates and be more severely impacted by EAB than trees in managed landscapes.
Finally, the Cipollinis fear that a close relative, pygmy fringetree, Chionanthus pygmaeus, is at particularly high risk because it is endemic to only a few counties in the sandhills of central Florida. The species is already classified as endangered by both the state and the federal governments. The pygmy fringetree is smaller than white fringetree, so its size might help it escape attack. However, adults achieve sizes comparable to that of fringetree in some cases. So when EAB reaches Florida, the specie appears to be highly vulnerable.
SOURCES
Cipollini, D. and K. Cipollini. 2026 The Fate of a Wild White Fringetree (Chionanthus virginicus) Population in Ohio 10 Years After Invasion by Emerald Ash Borer (Agrilus planipennis) Forests 2026, 17, 712
Deschênes, É., C.J.K. MacQuarrie, L. Scott, C. Zimmerman, and I. Aubin. 2026. Ash population dynamics after two decades of emerald ash borer infestations in Canada. Canadian Journal of Forest Research. Can. J. For.Res. 56: 1–13 (2026) | dx.doi.org/10.1139/cjfr-2026-0075
Wilson, C.J, L. Labbate, T.R. Petrice, T.M. Poland, D.G. McCullough. 2025. Ongoing regeneration of ash and co-occurring species 20 years following invasion by emerald ash borer. Forest Ecology and Management 580 (2025) 122546
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Hernández‐Gutiérrez, Nichols, and Kelly (2026) (full citation at the end of the blog) analyzed interactions between this genus of beetles and oaks (Quercus species). In this context, they report that 32 Agrilus species from Africa, Asia, Europe, and North and South America use 51 Quercus species as hosts in their native range. Eighteen (56.25%) use only Quercus hosts.
Oaks that host Agrilus species cluster in several clades, i.e., the entire Section Cerris and one clade of Section Ilex and two clades of section Lobatae. Clades where Agrilus hosts are underrepresented are Sections Cyclobalanopsis and Quercus &, surprisingly, two clades in Section Lobatae.
Their analysis indicated that beetle-oak interactions involving all the 32 Agrilus species and 105 Quercus species in their study had a significant likelihood of being damaging. For example, northern red oak (Q. rubra) is already known to host six Agrilus species. Hernández‐Gutiérrez, Nichols, and Kelly (2026)’s analysis indicated that this tree species might be utilized by all 32 Agrilus species assessed. The tree’s wide distribution (both native and introduced) places it close to other known or probable hosts, which exacerbates the risk of an interaction. Another 26 Quercus species are predicted to host ten or more Agrilus species.
The model was not successful in predicting hosts of A. auroguttatus (goldspotted oakborer) in its introduced range in California.
black oak (Quercus kelloggii) in Cleveland National Forest killed by GSOB; photo by F.T. Campbell
It also predicted that few of China’s oak species might host Agrilus beetles. However, as Dr. Robert Haack notes (pers. comm. June 2026), larval hosts have been identified for only 13% of the nearly 1,200 Asian Agrilus.
Some Agrilus species have a very large number of predicted novel interactions. A. graminis and A. angustulus are predicted to have more than 40 novel oak hosts. Both have numerous known oak hosts; so their “polyphagous” nature is already documented.
Since two-thirds of 666 Agrilus species with known larval hosts exploit only one plant genus as a host in their native range, Hernández‐Gutiérrez, Nichols, and Kelly (2026) assert that they might spread faster if introduced to homogeneous rather than species-rich habitats. Dr. Haack believes that this statement is too broad (pers. comm. June 2026).
A. bilineatus; photo by Christina Butler via Wikimedia
Twolined chestnut borer
Haack and Blank (2024) document that the twolined chestnut borer, Agrilus bilineatus has preferentially infested apparently healthy English oak (Quercus robur) trees over healthy native oak trees in Michigan. In North America, A. bilineatus is a major pest of oaks and American chestnut (Castanea dentata)when they are stressed by drought or other factors. Infestation typically begins in the upper crown and proceeds downward; tree death often occurs within three years.
At several sites in southern Michigan, where English oaks were intermixed with native white oaks (Quercus alba) and close to northern red oaks (Quercus rubra), A. bilineatus preferentially infested Q. robur trees that initially appeared healthy. Further study revealed that many of the English oaks attacked had low or depleted root starch levels.
Although these findings are cause for alarm, English oak is already used by 11 species of Agrilus in Europe. Perhaps the tree and ecosystem might have sufficient defenses in Europe. Meanwhile, A. bilineatus has been reported in Turkey as of 2018; I have found no recent information about the damage whether it is causing any damage there.
Data from Britain and Worldwide
Peyton et al. (2026) reviewed the effectiveness of a “horizon scanning” exercise conducted for Great Britain (England, Scotland and Wales). They report that 3,248 recognized non-native species have been detected in Great Britain, of which 2,016 have established self-sustaining populations. Some 194 (~10%) are considered invasive, that is, have negative impacts on biodiversity and wider ecosystem viability. These consist of 108 terrestrial species, 47 freshwater species, and 39 marine species. These bioinvaders cost the British economy an estimated ~£3.9 billion per year (the bulk of the damage is attributed to ash dieback, caused by the fungus Hymenoscyphus fraxineus).
In the decade between completion of the “horizon scan” and the present, 143 species were recorded as being introduced. The horizon scan predicted 31 of these species, 22%. Peyton et al. (2026) consider this to be success.
Peyton et al. (2026) report that globally, ~ 6% of non-native plant species are ranked as invasive. Among invertebrates, this proportion rises to 22%. Considering vertebrates introduced to Europe or North America, the figure is more than 50%!! I welcome global data that support my call for rethinking the “rule of tens” long relied on for estimating the proportion of non-native species that are invasive.
Discussing bioinvaders’ role in causing extinctions, Peyton et al. (2026) report that 30 predators have been linked to declines and extinctions of 738 vertebrate species.
Peyton et al. (2026) also discuss the difficulty in predicting an introduced species’ impacts when in some cases the time lag between introduction and presence in the wild or between establishment and spread w/in the region can last decades or even a century. They cite as an example Senecio squalidus, which escaped the Oxford Botanic Gardens in the 1700s but started to spread only during mid-1900s.
Australia
More than 300 non-native insect pests, pathogens and nematodes have established on tree or shrub hosts in Australia; 20% have caused moderate to high impacts to commercial plantations, urban forests, or trees in natural ecosystems (Carnegie et al. 2026).The rate at which non-native forest pests and pathogens have been detected in Australia has doubled since 2018 compared to earlier decades: from ~ 1.5 to ~ 3 per year. Carnegie et al. (2026) attribute this rise to greatly expanded official surveillance efforts. Still, three-quarters of the most recent detections came too late for eradication to be attempted.
The Forestwatch program (inaugurated – under a different name – in 2022) includes pathogens. I rejoice!!! Still, the target species threaten primarily tree species not native to Australia but important to commercial forestry or urban forests: Asian longhorned beetle, burnt pine longicorn (Arhopalus ferus), pine pitch canker, pine wilt disease, red turpentine beetle (Dendroctonus valens), Asian spongy moth, red needle cast, and sudden oak death. The exceptions are strains of Austropuccinia psidiinot yet intro to Australia, and eucalypt leaf blight (caused by Teratosphaeria destructans) (Carnegie et al. 2026).
Austropuccinia psidii infection on Melaleuca in Australia; photo by John Tann via Flickr
Among the introduced pests causing the greatest damage to native species are
Phytophthora cinnamomi: this soil fungus can kill 40% of the plant species in the southern portion of Western Australia – which is one of 36 “Biodiversity Hotspots” recognized by the Critical Ecosystem Partnership Fund.
Austropuccinia psidii (cause of myrtle rust) in natural ecosystems; Members of the host family Myrtaceae occur in 11 of 13 major vegetation formations on Australia. Various authorities have identified 76 species as at risk to the rust.
I hope the Australians are developing strategies for landowners to counter damage by the polyphagous shot hole borer (Euwallacea fornicatus) and its associated fungus (Fusarium euwallaceae). DMF Outbreak detected near Perth, Western Australia, in 2021 – apparently three years after the actual introduction. By June 2025 authorities had determined that it was too widespread to be eradicated, so landowners will be responsible for any management. (Carnegie et al. 2026) Impact is predicted to be greatest in urban landscapes, and cost up to AU$9.7 M per annum to manage.
Phytophthora pluvivora was first detected in Australia on an English oak, Quercus robur. However, it has since been recorded on native species in the Blue Mountains, including the critically endangered dwarf mountain pine (Pherosphaera fitzgeraldii) in a National Park.
SOURCES
Carnegie. A.J., B.A. Summerell, C. Trollip, F. Tovar, D.I. Smith, and J. McDonald. 2026. Sentinel trees for early detection of non-native forest pests and pathogens in Australia. Front. For. Glob. Change 9:1801183. doi: 10.3389/ffgc.2026.1801183
Haack, R.A. and R.B. Blank. 2025. Susceptibility of English Oak (Quercus robur) to the Twolined Chestnut Borer, Agrilus bilineatus (Coleoptera: Buprestidae): Observations from Michigan. The Great Lakes Entomologist. 57: 113-125. https://doi.org/10.22543/0090-0222.2492
Hernández‐Gutiérrez, E., R.A. Nichols, and L.J. Kelly. 2026. Combined phylogenetic and geographic data can predict plant–pest interactions with high accuracy. New Phytologist (2026) doi: 10.1111/nph.71306
Peyton, J.M., S. Rorke, D.C. Aldridge, O.L. Pescott, K. Dehnen- Schmutz, D.G. Noble, J. Sewell, A.J.A. Stewart, T. Adriaens, B.C. Beckmann, J. R. Britton, J. Brodie1, P.M.J. Brown, I.C.N. Cavadino, P.F. Clark, A.M. Dunn, J.Foster, C. Harrower, M.C. Harvey, M.C. Jackson, T. Jones, C.A. Maggs, G. Martin, F. Mathews, A.C. Mill, D. Murphy, E. Paganini, R. Payne, W. Rabitsch, T. Renals, K. Schönrogge, R.H. Shaw, G.C. Smith, P.D. Stebbing, P.A. Stroh, H. Tidbury, E. Tricarico, J. Vallet, K.J. Walker, L.E. Wood, C.A. Wood, B. Woodcock, H.E. Roy. 2026. Assessing the success of a horizon scanning approach in predicting invasive non- native species arrival. J Appl Ecol. 2026;63: https://doi.org/10.1111/1365-2664.70217
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Pananax ship; photo by 2005emsaguinden via Wikimedia
Imports from Asia, specifically China, had surged in the first half of 2025 as importers tried to get ahead of new tariffs. Blog They then fell sharply, although there was also a shift in Asian suppliers from China to Vietnam and India. At the end of 2025, experts expected the slump to persist into 2026. However, now we learn that U.S. imports from Asia jumped 13% from April to May, to 1.68 million TEUs (Michael Angell, Journal of Commerce, June 16, 2026). According to Politico, the shift to importing goods from Vietnam, and to a lesser extent to Thailand and Indonesia, has continued.
American cities continue to compete for to build larger ports so they can receive more cargo. Of course, more cargo means more incoming containers. As my blogs document (click “wood packaging” in the “Categories” section below the “Archives”), containers and especially the crates and pallets inside them too often transport wood-boring insects and other invasive species.
My source of information on these developments is from the Journal of Commerce website. Unfortunately, access is blocked by a paywall.
Recent reports concern port cities dredging and widening their ports in hope of attracting increased import volumes.
Norfolk; photo by PghPhxNfK via Wikimedia
Norfolk, Virginia
This month the Virginia Port Authority (VPA), Governor Abigail Spanberger and other federal, state, and local officials inaugurated the Port of Norfolk’s expanded capacity. At 55 feet, Norfolk is now the deepest and widest port along the East Coast. The project, begun in 2019, will allow Norfolk to handle vessels carrying up to 18,000 TEUs [twenty-foot equivalent containers]. Authorities are encouraging ocean carriers to call there.
The port was visited by 1,497 container ships over the past 12 months; nearly a quarter had a capacity greater than 11,000 TEUs.
Virginia’s container volumes grew on average 3% annually between 2017 and 2024. In 2025, total container volume fell 8% from 2024 because shippers hoped to avoid tariff deadlines by unloading cargo at a first or second port of call. Virginia was the six-busiest US port for imports this year through May, handling 635,205 TEUs. That was up 2.6% from the same five-month period a year ago. To attract shippers to Norfolk as a first or even second port of call for more container services, Virginia Port Authority CEO Sarah McCoy said, Virginia needs to emphasize rail links. This involves expanding the region’s distribution center network.
The project to dredge and widen the port began in 2019. The total project has a budget of $1.4 billion. The project included adding two ultra-large container vessel (ULCV) berths at each of its two container terminals. The final component is to renovate the North Berth, including developing a fifth ULCV berth, plus four new ship-to-shore cranes and semi-automated stacking cranes in its container yard. Completion is scheduled for mid-2027. At that time, the port’s total handling will reach 5.8 million TEUs. The International Longshoremen’s Association (ILA) is contesting installation of semi-automated stacking cranes in a series of lawsuits.
Other East Coast ports are also adding terminal capacity. This includes the Port of Baltimore, where a shipping company is developing a container terminal that includes handling two ULCVs simultaneously. Apparently they are not deterred by the months-long closure of the Port of Baltimore caused by a ship colliding with a bridge downstream from the port.
[Information from Michael Angell, Senior Editor, East Coast Ports. “Norfolk aims to be preferred USEC gateway with deeper port” June 18, 2026]
U.S. Army Corps of Engineers dredge in the Savannah River
The Port of Savannah remains dominant. In 2025 Savannah and the roll on-roll of facility in Brunswick together handled nearly 5.7 million TEUs, an increase of 2.6% or 146,000 TEUs compared to 2024. Even so, Georgia Ports is financing a $4.5 billion port and inland infrastructure plan which will add five new container berths in Savannah, the most new berths of any U.S. port and one new RoRo berth in Brunswick. Port officials brag about their rapid turnover of cargo from ship to rail. Forty-two double-stack trains per week take this cargo to Atlanta, Memphis, Nashville, Charlotte and Orlando. The Port Authority also operates two inland terminals – in in Chatsworth and Gainesville, Ga.
Corpus Christi
As I have reported in previous blogs, Gulf Coast ports are also upgrading to receive more shipments. DP World, a Dubai-based terminal operator, is negotiating to develop a container terminal at the Port of Corpus Christi. This would open this port to container logistics.
Corpus Christi is already the third-busiest port in the US by gross tonnage, behind Houston and South Louisiana. The bulk of cargo is petroleum and bulk commodities. (In past years, dunnage associated with bulk commodities proved to cause pest-infestation problems in nearby Houston.)
dunnage piled on the dock at Houston; photo by S. Useman
Last year Corpus Christi completed an eight-year, $625 million project to widen and deepen its main shipping channel (from 47 feet to 54 feet). The expansion allows visits by super-post-Panamax container vessels.
[Information from Michael Angell, Senior Editor, East Coast Ports. “DP World begins talks with Corpus Christi for container terminal project” June 16, 2026]
Warehouse Capacity
Leaders of logistics managers note that importers and domestic manufacturers are diversifying their supply chains – both warehouse facilities and shipping routes – because of ongoing supply chain disruptions even years after COVID 19. While many industrial warehouses across the U.S. still have vacancies (averaging above 7%), some markets are filling up. They mention Chicago, Indianapolis, Memphis, Dallas, and Kansas City. These cities’ warehouse vacancies average ~5%. Indianapolis leads; asking rents for industrial space there have climbed more than 50% over the past five years. Phoenix also anticipates increased demand for space, although its current vacancy rate is 10.6% – the highest vacancy rate of the 25 largest markets.
The move away from Los Angeles-Long Beach began years ago. To move freight inland, shippers need options, particularly access to intermodal rail.
shipping containers at Long Beach in the early 2000s; photo courtesy of Bob Kanter of the Port of Long Beach
The experts attribute this growth to the need for more flexibile supply chains and demand for materials used in constructing data centers. Demand for the second category is reportedly strongest in the Southeast, followed by interior central markets.
[Information from William B. Cassidy, Senior Editor, Trucking. “US warehousing expanding faster at key inland hubs” June 18, 2026]
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
ash trees (Fraxinus spp.) killed by EAB along Mattawoman Creek, Prince Georges County, Maryland; photo by Leslie A. Brice
I report here on action so far on funding vital agencies in Fiscal Year 2047 (which begins on October 1, 2026). I describe the various programs roles in addressing the invasive species threat briefly in my earlier blog.
USDA Forest Service
The House Appropriations Committee approved the FY27 Interior Appropriations bill (which includes the USFS) in early June. I greatly appreciate Congress’ continued support for two USFS programs that are vitally important in protecting resilience of the Nation’s forests in the face of invasions by non-native pests and plants: Research and Development and Forest Health Management (in the State, Private, and Tribal lands division). The Trump Administration had – for the second year in a row – called for ending these programs’ specific appropriations.
Research and Development
In my testimony I had asked the Subcommittee to maintain funding for R&D at the Fiscal Year 2026 Continuing Resolution level of $308 million. Instead the Subcommittee ignored the Administration’s request and provided $295 million – a fairly small reduction under the circumstances. Funding for the Forest Inventory and Analysis (FIA) program continues to see small increases — to $34 million.
While there is no specific line item for invasive species in the R&D budget, the report does encourage the USFS to address high priority invasive species, pests, and diseases, including the emerald ash borer, an unspecified “bark beetle”, spotted lanternfly, and Spruce Budworm. The list also includes three invasive plant species: buckthorn, Amur honeysuckle, and Callery pear. This language is not tied to a specific program, so it is unclear what will actually result.
In the R&D section of the report, the Committee “recognizes the significant damage invasive species can cause throughout forests, including urban forests, and encourages the USFS to continue reforestation efforts, including through the management of woody invasive species & tree planting in urban areas.” Again, the wording seems somewhat misplaced since the R&D program does not carry out tree planting.
State, Private, and Tribal forests
I had asked the Interior Appropriations Subcommittee to maintain funding for State, Private, and Tribal forests program at the FY26 Continuing Resolution level of $310.6 million. Again, appropriators ignored the Administration’s request and provided $283,500,000 – a fairly small reduction. The Forest Health Management program was funded at $58 – of which $16 million is specified for efforts on federal lands, $42 million for helping state and tribal agencies and private landowners to manage pests on their lands.
The Subcommittee report emphasized the importance of working with Colorado to curtail spread of mountain pine beetle d associated wildfire risk. Music to my ears is the Committee’s statement encouraging the USFS to work with state & local agencies to counter the high rate of tree mortality due to the goldspotted oak borer infestation in Southern California. It advised prioritizing insect research, prevention, suppression, & mitigation projects that support community wildfire protection & State forest action plans. Since none of the members of the House Interior Appropriations Subcommittee is from California, I am pleasantly puzzled.
coast live oak (Quercus agrifolia) killed by goldspotted oak borer; Heisey County Park, San Diego County, California. Photo by F.T. Campbell
The Interior Appropriations Subcommittee expressed continued concern about poor regeneration of eastern white oak (Quercus alba). (Of course, several other tree species are also regenerating poorly but a strong lobby is tied to oaks due to their economic importance.
i’iwi – a formerly common Hawaiian honeycreeper badly suppressed by avian malaria; photo by by James Petruzzi
Hawaii’s endangered birds
I am very pleased that the House Interior Appropriations Committee has provided $1,250,000 is for continuing research to protect Hawaiian forest birds from the dire extinction threat arising from non-native mosquitoes carrying avian malaria & other pathogens.
USDA Animal and Plant Health Inspection Service
APHIS is responsible for preventing the introduction and spread of pests and invasive plants that harm agriculture, including forests. APHIS policy guides port inspections carried out by the DHS CBP. APHIS inspects imported live plants. Unlike the USFS, APHIS has the support of the Trump Administration, so funding levels have remained steady. Of course, continuing introductions of new pests and spread of established ones – and inflation – have increased the cost of countering invasions, so the agency continues to fall behind despite its relatively privileged position.
The House of Representatives adopted the FY27 Agriculture Appropriations Bill in early June. I apologize for the obscurity of the FY26 funding levels. Figures are in millions of dollars.
FY2025 enacted FY27
APHIS total $1,148 $1,157
Plant health subtotal $387.5 $387.6
Agric. quarantine $35.5 $35.5
Field crop and rangeland $12 $10
Pest detection $29 $29
Methods development $21.5 $21
Specialty crops $206.5 $209
Tree and wood pests $59 $58.6
In its report, the Subcommittee on Agriculture Appropriations did not express concern about the issues that I had raised in my testimony. Instead, it mentioned several agricultural pests, e.g., citrus greening, fruit flies, a non-native beetle threatening nut orchards, and spotted lanternfly (Lycorma delicatula). The Subcommittee also urged APHIS to work with the USDA Forest Service to counter the spread of two native wood-borers, mountain pine beetle (Dendroctonus ponderosae) in Colorado and southern pine beetle (Dendroctonus frontalis). Since both are native, APHIS’ role is unclear. The Subcommittee mentioned two invasive plants: glossy buckthorn (Frangula alnus) – saying it threatens biodiversity, especially in the Allegheny National Forest in Pennsylvania; and Carizzo cane (Arundo donax), saying it contributes to tick invasions in addition to hampering detection of human immigrants along the Rio Grande.
sounder graphic by Jack Mayer, Savannah River National Laboratory
The bill increases funding for the national feral swine management program by $500,000. (I cannot determine total the appropriation). The Agriculture Appropriations Subcommittee had urged APHIS Wildlife Services to prioritize states with the largest invasive populations – although this choice contradicts standard advice for managing invasive species, including feral swine.
In one action linked to our concerns about wood-borer introductions via wood packaging, the Subcommittee encouraged APHIS to recognize sulfuryl fluoride as a treatment for logs, wood products, & wood packaging. This fumigant has been accepted under the international (ISPM#15) since 2013.
wooden pallets discarded next to wooded border in Glacier National Park, Montana; photo by F.T. Campbell
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
eastern (Canadian) hemlocks in Nova Scotia killed by hemlock woolly adelgid; photo by Celia Boone, NSDLF
Two recent studies show that climate change is driving changes to pest ranges. These pose a serious threat to forests of eastern Canada.
A study by Aubin et al. (2026; full citation at end of blog) assessed the risk that climate change would exacerbate the impacts of 14 non-native forest insect pests The 14 insect species were selected from an original group of 76 species using the following criteria:
1) They have the potential to cause mature tree mortality to at least one of the 37 tree species most abundant in Canada; and
2) They have been detected in Canada recently or have established populations actively spreading through the Canadian landscape.
Major Findings
Twenty-four tree species are affected currently by at least one of the 14 non-native insect pests (excluding Asian longhorned beetle; see below). (I list the tree and insect species below.) Four of them are already considered globally endangered or threatened due to invasive pests: black and white ash (Fraxinus nigra and F. americana), whitebark pine (Pinus albicaulis) and eastern hemlock (Tsuga canadensis). Another ash species (blue ash; Fraxinus quadrangulata) and another pine (limber pine; Pinus flexilis) are considered rare or threatened species in Canada.
brown spruce longhorned beetle; photo by Udo Schmidt via WikiMedia
Of the 24 species at risk, black spruce (Picea mariana) is most exposed because it grows in areas where eastern spruce gall adelgid and brown spruce longhorned beetle (Tetropium fuscum) are established. Expansion of these two pests could potentially reach 75% of black spruce biomass in Canada. However, Aubin et al. (2026) expect tree mortality to be limited because these insects target trees that are already stressed or weakened. Of course, the changing climate might increase the trees’ susceptibility. Thirty percent of the black spruce’s range is projected to be outside its current climatic niche by 2040.
The 24 tree species currently affected by at least one of the 14 non-native insects (excluding ALB) collectively constitute 3.2 billion tons of tree biomass. The impact is projected to increase more than four-fold — to 13.6 B tons of biomass — within two decades. Reinvasion by the Asian longhorned beetle would put at risk six additional tree species in the genera Acer, Betula, and Populus. Their jeopardy would add another 3.1 billion tons of live tree biomass to the “at risk” category.
Aubin et al. (2026) note that affected trees might remain alive but moribund, that is, lose their functional role within the ecosystem. for several years before finally dying. Therefore their analysis might underestimate pests’ impact on the forest. Their example is American beech – as weakened by beech bark disease. A reminder: beech bark disease causes widespread death of mature beech – opening the canopy and eliminating such wildlife-supporting aspects as nesting cavities and abundant crops of nuts. Vigorous root sprouting results in dense stands of young beech, crowding out other species.
Hotspots of greatest vulnerability
The analysis identified two hotspots of greatest vulnerability: north-central British Columbia near the border with Alberta, and along the St. Lawrence Seaway near the border with the United States. The pest pressures differ.
The threat to the British Columbia hotspot comes from expansion of mortality in dense pine forests caused by the native mountain pine beetle (Dendroctonus ponderosae). Aubin et al. (2026) describe a two-pronged impact from the changing climate: trees lose vigor because they become maladapted to the new growing conditions (temperature and drought); while the beetle increases the frequency of outbreak due to reduced overwintering mortality.
Along the St. Lawrence Seaway (southern Ontario and Quebec) the threat comes from a suite of non-native insects, including emerald ash borer, hemlock woolly adelgid, and beech scale. While emerald ash borer and beech scale were introduced directly to Canada by international trade, hemlock woolly adelgid spread across the border from the U.S. Further northward expansion of all three is projected under both low and high emission climate change scenarios.
beech bark disease in New Hampshire; photo by Eli Sagor via Flickr
Most alarming is that some regions in eastern Canada are vulnerable to invasion by all 14 insect species. Two additional pests loom: Aubin et al. (2026) fear northward expansion might reintroduce the Asian longhorned beetle or introduce the southern pine beetle (Dendroctonus frontalis). The latter has been expanding northward in the U.S.
Pests often move across the Canada-U.S. border. In addition to the five pests mentioned above, spongy moth, and two pathogens, the beech leaf disease nematode, and the oak wilt fungus, have spread from the U.S. into neighboring parts of Canada. The woodwasp Sirex noctilliowas probably introduced simultaneously to both countries. Winter moth and beech bark disease spread from Canada to the U.S. I worry that the brown spruce longhorned beetle might do the same.
Asian longhorned beetle: can this disaster be averted?
As noted above, the Canadians are alarmed by the prospect that the Asian longhorned beetle might be reintroduced – either by spread from extant populations in the United States or directly on imports from China. Aubin et al. (2026) note that the two earlier – successful! — eradication programs were expensive, costing an estimated CND$35.5 million. This expenditure is dwarfed by the costs estimated to arise from an unmanaged invasion: CDN$431 million annually in timber products and CDN$358 million annually in edible maple products. There would also be enormous ecological impacts, including threats to an additional 3,08 metric tonnes of tree biomass comprising ~24.96 tons of CO2 equivalent.
Canada’s central boreal forest is at lower risk both in terms of exposed tree biomass and number of invasive insect species present. The reasons are not understood. Aubin et al. (2026) suggest that the boreal ecosystem is more resistant to invasion due to a combination of environmental barriers and native natural enemies. For example, the introduced woodwasp Sirex noctilio did not cause widespread pine mortality in the region, probably due to antagonistic interactions with other subcortical species.
Although it does not fit this definition, Aubin et al. (2026) also evaluated the Asian longhorned beetle (ALB Anoplophora glabripennis), because of its huge impact if it is reintroduced to Canada (see above).
One of the focal groups, beech scale, Cryptococcus fagisuga, differs from the others because it is a vector of a tree-killing fungal pathogen (Neonectria coccinea); the scale itself does not cause notable harm.
Aubin et al. (2026) acknowledge that additional species represent a possible threat to Canadian forests. Therefore their study does not represent the total risk posed by all potential invasive insects in Canada, but provides a snapshot of selected, current vulnerabilities.
The 37 most abundant tree species in Canada together represent 88% of total mature forest tree biomass in Canada. They include 17 conifers and 7 deciduous trees:
Abies amabilis, Ab. balsamea, Ab. lasiocarpa; Acer rubrum, Ac. saccharinum, Ac. saccharum; Alnus rubra; Betula alleghaniensis, B. papyrifera; Callitropsis nootkatensis; Fagus grandifolia; Fraxinus american, F. nigra; Larix laricina, L. occidentalis; Picea engelmanii, P. glauca, P. mariana, P. rubens, P. sitchensis; Pinus albicaulis, P. banksiana, P. contorta, P. ponderosa, P. resinosa, P. strobus; Populus balsamifera, P. grandidentata, P. tremuloides; Pseudotsuga menziesii; Quercus rubra; Thuja occidentalis, T. plicata; Tilia Americana; Tsuga canadensis, T. heterophylla, T. mertensiana
The 14 insects collectively have 63 host tree species in Canada. The pine genus is susceptible to the largest number of pests. Genera found to be not vulnerable to any of the 14 insects are Acer, Callitropsis, Populus, Pseudotsuga, Tilia, and Thuja. The proportion of total exposed tree biomass in Canada varied by species, from 8% of Jack pine (Pinus banksiana) to 95% for red spruce (Picea rubens).
red spruce in West Virginia; photo by Famartin via WikiMedia
Although most of the 14 insect species are projected to benefit from larger areas of suitable climate in Canada over the next 20 years, there are interesting exceptions: European oak borer (Agrilus sulcicollis), brown spruce longhorned beetle, and winter moth (Operophtera brumata). Distributions of the host tree species are projected to change insignificantly over the 20 years covered by the study.
With northward expansion of suitable climates for 12 of the 14 species, large areas of the boreal forest will be exposed to potential invasion. The entire Canadian distribution of 13 of the 37 dominant tree species might be at risk: three Abies, two Betula, both Fraxinus, Picea mariana, Pinus contorta, Picea glauca, Pinus banksiana. Aubin et al. (2026) mention specifically Engelmann spruce (Picea engelmanii) and American beech (Fagus grandifolia).
Other Factors
The analysis did not consider possible alterations of the insects’ life history traits other than potential expansion of their distributions. Warmer temperatures can cause changes in voltinism, diapause periods, development rates, reproduction, and population growth; cumulatively, these changes might alter their invasion dynamics. The mountain pine beetle is an example. Populations have experienced outbreaks more frequently, so increasing the species’ invasion threat and severity. On the other hand, shifts in temperature and precipitation could decouple the phenology of trees and their associated pests, reducing insect survival. Finally, complex changes in tree tissue and their secondary defensive metabolites (see below) also could alter interactions between non-native insects and their new hosts – possibly exacerbating or mitigating the herbivores’ impacts.
Aubin et al. (2026) remind us that loss of a dominant species might lead to reorganization of forest composition and structure. They expect the impacts to be particularly critical in stands with low tree diversity, such as the pure jack pine stands in the eastern boreal forest. Loss of a foundation species might also profoundly disrupt ecosystem functions, carbon budgets, wildlife habitats and stand productivity. They cite cascading effect on aquatic invertebrate communities and invasions by non-native plant species following widespread death of ash trees caused by the emerald ash borer. Finally, death of some species reduces functional redundancy within tree communities, and a shrinking pool of viable native replacement species. The widespread planting of ash trees in urban areas after the demise of most elms is such a case.
American elms in Jeanne d’arc Park, Quebec; photo by Cephus via Wikimedia
The possibility that the tree hosts might increase production of defensive metabolites was corroborated by Mike Aucott in a different context. Dr. Aucott is retired from the New Jersey Department of Environmental Protection. He authored a guest blog in December 2022, in which he discussed changes in plant chemistry brought on by the 50% increase in atmospheric CO2 levels over the last century. By happenstance, Dr. Aucott engaged in an exchange of letters in Science (2/26/26), in which he reiterated the likelihood that plants, “fertilized” by access to this nutrient, might be better able to fend off insect attacks. (See the “Sources” section for references to additional information on this phenomenon.)
Compounding Threat: The Spruce Budworm in Eastern Canada
The spruce trees of eastern Canada face another pest threat: the native spruce budworm (SBW, Choristoneura fumiferana). It is already a major defoliator in North American boreal forests.
Boulanger et al. (2025; full reference at end of this blog) documented pronounced changes in the moth’s range, especially in the East. Over the past 60 years, suitable climate conditions for the budworm have expanded northward. On the other hand, winter mortality has increased in southern parts of its range due to warmer temps. Overall, the total area highly suitable for population growth remained virtually the same. Still, the budworm’s earlier activation might exacerbate its impact on the previously less vulnerable black spruce, Picea mariana. If so, this might fuel further increased population growth rates northward.
black spruce; photo by Laval University via WikiMedia
Like Aubin et al. (2026), Boulanger et al. (2025) found that Canadian forests in the east and Atlantic regions are likely to experience greater impacts on tree growth than are forests in western and central regions. The host most vulnerable to SBW, balsam fir (Abies balsamea), is a dominant or codominant species in the East and Atlantic regions. The fir is sparsely distributed in those areas of central and western Canada where the climate is becoming highly suitable for the insect. The frequent wildfires promote growth of young pioneer tree species, e.g., jack pine and trembling aspen, that do not support SBW.
At the most general level Boulanger et al. (2025) suggest that climate change might have already surpassed impacts of land use change on spruce budworm dynamics. Climate change puts additive and synergistic pressures on insects, which are already more sensitive than trees to climatic factors and able to adapt more quickly. As a result, climate change is becoming the most significant driver of recent declines in insect abundance and shifts in community structure, development, dispersal patterns, and phenology. (Again, see Aucott, above, for an alternative explanation.)
Boulanger et al. (2025) mention but do not discuss possible impacts of climate change and a shift in SBW distribution and tree hosts on a third trophic level, i.e., natural enemies. They note that many factors – not just climate suitability — influence trophic interactions. Another complication is that most SBW parasitoids require alternate hosts.
Boulanger et al. (2025) join others in urging forest managers to quickly adapt their management strategies to the novel climate-induced threats. They call for a proactive and integrated approach in forest management. Ecological research will be crucial to mitigate the compounded effects of climate change and to preserve the integrity & sustainability of forest ecosystems.
SOURCES
Aubin, I., A. Roe, B. Marquis, L. Boisvert-Marsh, J. Pedlar,S. Erni, B. Hamel, G. Lawrence, D. McKenney, T. Scarr. 2026. Vulnerability of Canadian forests to invasive insects under climate change. Accepted by the Canadian Journal of Forest Research.
Boulanger, Y., A. Desaint, V. Martel, M. Marchand, S. Massoda Tonye, R. Saint-Amant, et al. (2025) Recent climate change strongly impacted the population dynamic of a North American insect pest species. PLOS Clim 4(2): e0000488. https://doi.org/10.1371/journal. pclm.0000488
See also an article describing declining nutrient value of food crops in response to increased atmospheric C02 levels: https://www.washingtonpost.com/climate-environment/interactive/2026/carbon-pollution-diluting-key-nutrients-food/
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The Wildlife Society’s Invasive Species and the Forestry and Wildlife Working Groups are cosponsoring a workshop for the Lingering Hemlock Project. The workshop will be on Tuesday 28 April at 2:00 PM EDT.
The Lingering Hemlock Project is a subset of The Nature Conservancy’s “Tree Species in Peril” program. The project aims to locate and selectively breed eastern hemlocks with genetic resistance to the hemlock woolly adelgid (HWA).
Olivia Hall from the North Carolina Hemlock Restoration Initiative will share more about how natural areas can participate in the Lingering Hemlock Project. In the southeastern US, project partners can locate and record data on hemlocks that remain healthy despite HWA infestations. In the northeastern US, project partners can establish hemlock plots and monitor their health and decline annually.
Go here to learn more about the project & webinar, and find the link to join.
I have blogged about HWA often – although there is no simple method for finding the earlier blogs. In 2025 I posted 3 blogs – in March and one in August. If you need a reminder about HWA, visit TNC’s “don’t move firewood” website here.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm