Reminder: Friday is the deadline for commenting on APHIS’ draft environmental assessment for the Asian longhorned beetle eradication program in South Carolina. Comments should be submitted at https://beta.regulations.gov/commenton/APHIS-2020-0086-0001
The Center for Invasive Species Prevention submitted comments that supported the eradication effort because of the well-documented threat that the ALB poses to the forests of North America. We also supported the preferred alternative in the EA.
However, we found the environmental assessment (EA) to be deficient in several ways:
the EA does not identify the host species present in the program area – not even of the 5,800 trees inspected by the program as of mid-August.
the EA provides no estimate of the proportion of deciduous trees and shrubs in the area that are host species. Conifers dominate the area. This means that any fauna dependent on deciduous trees and shrubs for food and shelter already contend with limited resources. Consequently, while we concur with the EA that any impacts will be localized, they might be exacerbated by the relative rarity of hardwood species in the local area. It is particularly important that the EA address this question since the Programmatic EIS was written under the assumption that forests at risk to the ALB are like those in the Northeast and Midwest, where hardwoods dominate.
Without knowing the proportion of deciduous flora comprised of host species, no one can evaluate the amount of wildlife food that could be removed or treated by pesticides. Some wildlife species are potentially vulnerable, including those that feed on pollen and nectar (i.e., bees and other pollinators) and those that feed on insects and other invertebrates. The latter include two species listed federally as threatened species: the frosted flatwood salamander (Ambystoma cingulatum) and northern long-eared bat (Myotis septentrionalis). Also vulnerable are birds, 96% of which feed their young on insects and other invertebrates. I worry about sublethal effects and possible bioaccumulation. Aquatic organisms, especially invertebrates, might also be affected.
The information gaps in the EA highlight weaknesses in the Programmatic EIS, on which it relies. The most important gap is the dearth of pesticide dose/mortality data for terrestrial amphibians. Apparently, EPA has not required such studies before approving pesticides.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
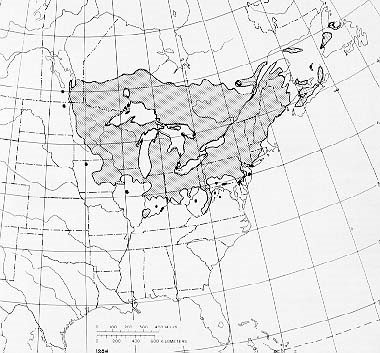
Another unique ecosystem being severely damaged by non-native tree-killing pests are the wetlands dominated by black ash (Fraxinus nigra). Black ash typically grows in fens, along streams, or in poorly drained areas that often are seasonally flooded. Such swamps stretch from Minnesota to Newfoundland; in the three states of Michigan, Wisconsin, and Minnesota, they cover a total of over 2 million hectares (Kolka et al. 2018).
Recent research allows us to understand the impending loss to these unique ecosystems that will be caused by the emerald ash borer (EAB).
Hydrology is the dominant factor that influences a host of ecosystem functions in black ash wetlands. Water levels are largely determined by a combination of precipitation and evapotranspiration rates. Black ash can thrive in wetter areas than most other tree species (Slesak et al. 2014). Water tables in these swamps are typically above the surface throughout early spring, followed by drawdown below the surface during the growing season with periodic rises following rain events. Water table drawdown coincides with peak evapotranspiration following black ash leaf out, demonstrating the fundamental control that this species has on animal and other plant communities (Kolka et al. 2018; Slesak et al. 2014).
Ecological Importance
Black ash generally dominate the canopy of these wetlands. Ash density can range from about 40% to almost 100%. Several other tree species are present, including northern white cedar (Thuja occidentalis), red maple (Acer rubrum), American elm (Ulmus americana) (Kolka et al. 2018), quaking aspen (Populus tremuloides), American basswood (Tilia americana), and bur oak (Quercus macrocarpa) (Slesak et al. 2014), balsam fir (Abies balsamea), balsam poplar (Populus balsamifera), and speckled alder (Alnus incana) (Youngquist et al. 2020). Black ash, by maintaining low water levels during the growing season, creates conditions under which these other trees can live but not thrive (summary of study by B.J. Palik, USDA Forest Service, here. Most other species lack the physiological adaptations of black ash or face pathogenic constraints (e.g., Dutch elm disease on American elm Ulmus americana) (Kolka et al. 2018).
Ash trees in these swamps are uneven-aged with canopy tree ages ranging from 130–232 years (Slesak et al. 2014). This complexity provides important habitat for many wildlife species, including ground beetle community assemblages (Kolka et al. 2018) and an abundance of aquatic macroinvertebrates. These are characterized and dominated by mollusks (Sphaeriidae, Lymnaeidae, Physidae), annelids (Lumbriculidae, Hirudinea), caddisflies (Limnephilidae, Leptoceridae), and dipterans (Chironomidae, Culicidae) (Youngquist et al. 2020).
a black ash swamp; source: Flickr
A major concern is that loss of trees – especially ash – might result in open marshes dominated by grasses, especially lake sedge (Carex lacustris). Conversion to sedge-dominated marshes has been observed in areas where trees have been removed as part of experiments to test various ecosystem responses to loss of the ash component (Slesak et al. 2014). Even if other trees took the place of ash, the substitutes might not support the same animal communities (see below).
Impact of Emerald ash borer and loss of black ash
Black ash is highly susceptibility to the EAB (Engelken and McCullough, 2020), so scientists expect severe impacts of the invasion in ash-dominated wetlands and – to a somewhat lesser extent — in forested stream systems’ riparian areas (Engelken and McCullough, 2020). They expect cascading impacts on 1) hydrology; 2) plant communities; 3) wildlife; 4) Native American cultures; and possibly even storage of carbon in vegetation and soils (Kolka et al. 2018).
1) Hydrology
Experiments suggest that loss of ash will cause higher water tables, especially during late summer and fall (Kolka et al 2018). This will result from reductions in evapotranspiration as large trees are replaced by shrubs and grasses (see below) (Kolka et al. 2018; Slesak et al. 2014). The higher water table might be exacerbated if higher annual precipitation levels predicted by climate change models occur. On the other hand, these models also predict a simultaneous increase in longer droughts, which might partially counteract higher precipitation and reduced evapotranspiration (Kolka et al. 2018). If they occur, these possible increases in drought length and frequency might enhance the establishment of less water-tolerant non-ash tree species in former black ash wetlands.
2) Plant Communities
Higher water tables are expected to reduce tree densities and promote conversion to open or shrub-dominated marshes. Several of the possible alternative tree species do not thrive as well as black ash under current conditions (Kolka et al. 2018). However, new hydrologic conditions might make forest restoration even more difficult because herbaceous plants transpire less water than trees, thus exacerbating the rising water tables (Slesak et al. 2014).
In upper Michigan, experiments which killed ash by cutting or girdling did not lead to an increase in growth rates of the remaining canopy species despite the increase in available resources (e.g., sunlight and nutrients) – presumably because of the raised water table (Kolka et al. 2081).
While some studies have found that black ash seedlings and saplings dominated the woody component of the swamp understory up to three years after ash were experimentally removed (Kolka et al. 2018), Engelken and McCullough (2020) found only eight saplings and a single seedling.
Scientists have planted several tree species in experiments to see which might be used to maintain the forested wetlands in the absence of black ash. The results are a confusing mix. Some species grew well once established – but had low levels of seedling establishment. Some trees planted on elevated microsites (hummocks) had the greatest survival and growth rates. (For specific data, see Kolka et al. 2018). A further consideration is tree species’ ability to adapt to warming temperatures already evident and expected to increase in coming decades (Slesak et al. 2014).
Consequently, Slesak et al. (2014) think it is likely that the EAB invasion will alter vegetation dynamics and cause a shift to an altered ecosystem state (e.g., open marsh condition) with higher water tables. They caution that the degree of ecosystem alteration will vary depending on site hydrology, annual precipitation, and period of time necessary for establishment of deeper rooted vegetation.
3) Wildlife
Moreover, any changes in vegetation will also affect the biota in more subtle ways through altered nutrient cycles. Black ash leaf litter is highly nutritious, having some of the highest nitrogen, phosphorus, and cation contents of any hardwood forest species (Kolka et al. 2018). Black ash leaves also decompose faster than most alternative tree species’ leaves (summary of Palik USDA Forest Service, here; Youngquist et al. 2018).
Youngquist et al. (2018) studied litter breakdown, litter nutritional quality, and growth of a representative invertebrate litter feeder – larvae of a shredding caddisfly (Limnephilus indivisus). They found that the larvae’s risk of death increased by a factor of three times or more when caddisflies were fed American elm, balsam poplar, or lake sedge leaves compared to black ash leaf litter. Even when the larvae lived – but matured more slowly because of the lower nutrition value of the leaves – they would still be vulnerable because they must reach metamorphosis before pond dry-down. In any planting done to maintain forested quality of wetlands, need to consider the nutritional quality of the leaf litter provided by replacements. Speckled alder was only apparently acceptable substitute; it was second to black ash in acceptability to caddisflies (Youngquist et al. 2020)
In fact, Youngquist et al. (2020) concluded that plant and detritivore biodiversity loss due to EAB invasion could alter productivity and decomposition at rates comparable to other anthropogenic stressors (e.g., climate change, nutrient pollution, acidification). The result will be altered biogeochemical cycles, resource availability, and plant and animal communities.
Scientists are also concerned about the impact of ash tree mortality on forest connectivity. Conversion of wooded swamps to shrub-and sedge-dominated wetlands will result in the loss of important micro-habitats that are already limited across the forested landscape and may also reduce availability of critical habitat for migrating birds. These changes will exacerbate on-going changes in land use in the Great Lakes region that are causing loss of forest habitat and forest homogenization. As yet, the magnitude of the impact on wildlife is unclear (Kolka et al. 2018).
black ash baskets – displayed at 2006 conference photo by Faith Campbell
4) Cultural importance – baskets
Native Americans living in the range of black ash have utilized the wood to make baskets and other tools for thousands of years. Baskets had numerous uses, such as packs for carrying items, fish traps, and for preparing food and storing household items. Ash items also had ceremonial uses and they are highly sought as gifts and in trade. The skill needed to select a good tree and work the wood is handed down through the generations and is an important part of tribes’ culture (Benedict 2010).
Discussion of these cultural traditions can be found as Powerpoints here and here.
Concerned by the spread of EAB and probable impact on black ash swamps, the USDA Forest Service has initiated major research studies with the goal of filling in the numerous knowledge gaps and developing management recommendations. A large-scale study using various manipulations to simulate the EAB invasion was initiated in the Chippewa National Forest in northern Minnesota in 2009. A companion study began in the Ottawa National Forest in Michigan in 2010 (Kolka et al. 2018). The Slesak, Youngquist, and Kolka publications cited in this blog report results of some of the studies in this project. Other studies of black ash conditions, including regeneration, at various stages of the EAB invasion wave are being carried out by Deb McCullough, Nate Siegert, and others. They are working at sites from Michigan to New England (D.G. McCullough, pers. comm.).
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report here.
For a great discussion of black ash basketweavers, see Anne Bolen, A Silent Killer: Black Ash Basket Makers are Battling a Voracious Beetle to Keep their Heritage Alive, American Indian Magazine, Spring 2020, available here.
Engelken, P.J. and D.G McCullough. 2020. Riparian Forest Conditions Along Three Northern Michigan Rivers Following Emerald Ash Borer Invasion. Canadian Journal of Forest Research. Submitted
Kolka, R.K., A.W. D’Amato, J.W. Wagenbrenner, R.A. Slesak, T.G. Pypker, M.B. Youngquist, A.R. Grinde and B.J. Palik. 2018. Review of Ecosystem Level Impacts of Emerald Ash Borer on Black Ash Wetlands: What Does the Future Hold? Forests 2018, 9, 179; doi:10.3390/f9040179 www.mdpi.com/journal/forests
Slesak, R.A., C.F. Lenhart, K.N. Brooks, A.W. D’Amato, and B.J. Palik. 2014. Water table response to harvesting and simulated emerald ash borer mortality in black ash wetlands in MN, USA. Can. J. Forestry. Res. 44:961-968.
Youngquist, M.B., C. Wiley, S.L. Eggert, A.W. D’Amato, B.J. Palik, & R.A. Slesak. 2020. Foundation Species Loss Affects Leaf Breakdown and Aquatic Invertebrate Resource Use in Black Ash Wetlands. Wetlands. Society of Wetland Scientists
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
We know the dire threats to Hawaiian forests from pathogens. Some threaten the most widespread tree – ohia. Others are insects threatening trees and shrubs in the remnant dryland forests.
The forests of smaller islands of the Pacific also appear to be facing severe threats – although I have been unable to find information on the current situation.
Guam and its Neighbors
The forests of Guam, Palau, and others in the Western Pacific are among those threatened.
They are geographically isolated and hard to reach, but that distance has not protected them from biological invaders. Their predicament illustrates the dominant role of global movement and trade in spreading pests. In this case, it’s mostly trade in ornamental plants.
These islands have unique flora and fauna. And true to invasive species experts’ expectations, they are vulnerable to bioinvaders. Guam’s most famous invasive species is the brown tree snake (Boiga irregularis), which over a few decades eradicated many bird species and the only native terrestrial mammal, the fruit bat.
Less known, but equally damaging, have been a group of insects that are decimating Guam’s native forest flora.
The most widespread arboreal species in the forests of Guam and neighboring islands is the Micronesian cycad, Cycas micronesica. Its range is Micronesia, the Marianas Group including Guam and Rota Islands; and several of the western Caroline Islands, e.g., Palau and Yap (Marler, Haynes, and Lindstrom 2010).
These forests have already absorbed severe habitat destruction as the sites of fierce fighting in World War II and – in some cases – construction of large military bases. Still, cycads were the most common species in the forest as late as 2002 (Moore, A., T. Marler, R. Miller, and L. Yudin. Date uncertain).
The Worst Pest: Asian Cycad Scale
The most severe current threat to the cycads are introduced insects, especially the Asian cycad scale Aulacaspis ysumatsui.
The cycad scale is native to Southeast Asia. It was first detected on Guam in 2003, when officials noticed that cycads planted near hotels had begun to die. However, this scale had already been spreading thanks to the trade in ornamental cycads. It was detected in Florida in 1996, on Hawai`i in 1998. It continued to spread rapidly in the western Pacific: to Rota in 2007, Palau in 2008 (University of Guam 2012). By late 2019, the scale had spread globally – numerous islands and neighboring mainland areas in the Caribbean (including Puerto Rico and US Virgin Islands), several US states in the Southeast, California, and Taiwan (Moore, Marler, Miller, and Yudin. Date uncertain.) and South Africa. (vanWilgen, et. al. 2020) Also, see the map prepared by CABI.
In every case, the scale has apparently been spread on nursery stock. It is difficult to contain by standard phytosanitary measures – visual inspection – because the scale is tiny and hides deep in the base of the plant’s stiff leaves and other crevices. (Marler and Moore 2010)
By 2005 the scale was killing the native cycad on Guam. Within four years, the millions of C. micronesica on Guam were reduced by more than 90% (Marler, T.E. and K.J. Niklas. 2011). The last time cycads on Guam reproduced in any significant number was in 2004 (Marler and Niklas 2018).
The severe impact of the scale was so rapid that the International Union for Conservation of Nature and Natural Resources (IUCN) changed its listing of C. micronesica from “near threatened” in 2003 to “endangered” in 2006. (IUCN Red List of Threatened Species Online 2008).
Scientists have made several attempts to introduce a biocontrol agent. However, the most promising – the lady beetle Rhyzobius lophanthae – has failed to control the scale, despite having become virtually ubiquitous on Guam. The beetle is too big to reach the significant proportion of scale insects living in small cracks and voids within the plant structures. Evidence from another cycad species indicates that the beetles also don’t prey on scale insects living beneath trichomes (fine hairlike structures on the leaves) or on parts of the plant close to the ground. (Moore, Marler, Miller, and Yudin. Date uncertain.).
Attempts to introduce a second biocontrol organism – the parasitoid wasp Aphytis lignanensis – were stymied by the presence of R. lophanthae (Moore, Marler, Miller, and Yudin. Date uncertain).
Micronesian cycad photo by Lauren Gutierrez
Other Invasive Species Attacking Cycads
The cycad blue butterfly (Chilades pandava) was detected in 2005 and spread throughout Guam within months (IUCN 2009). Also, it’s been found on Saipan (1996) and Rota (2006). The butterfly is native to southern Asia from Sri Lanka to Thailand and Indonesia. High populations can cause complete defoliation of new foliage. Repeated defoliations can kill the plant. Cycads on Guam are particularly vulnerable because the scale has already caused loss of most of their leaves. Butterfly larvae are often protected by ants (Anonymous).
On cultivated plants the butterfly can be controlled by microbial insecticides containing Bacillus thuringiensis kurstaki (Moore). Scientists at the University of Guam are exploring use of injected insecticides (Moore). They have found an egg parasite, but parasitism levels are low. Any biocontrol agent targetting larvae would have to contend with the ants (Anonymous).
A longhorned beetle (Dihammus (Acalolepta) marianarum) and a snail (Satsuma mercatorius) are also feeding on the cycads (Marler 2010).
The Indo-Malayan termite Schedorhinotermes longirostris was detected in 2011. The termites weaken the cycad stems, which are then toppled by feeding by introduced deer. The termites are also damaging the cycad’s reproductive structures (megastrobili). Termite attacks on cycads surprised scientists since cycads do not form true wood. The termite had probably been introduced recently because, as of 2011, it had been detected only near the Andersen Air Force Base airport (Marler, Yudin, and Moore 2011).
More Isolated – but Still Overrun
Scattered across the Pacific are groups of atolls, including Palmyra and Rose.
Despite their distance from other islands, they have all been visited by mariners for centuries. As a result, they have non-native species, including insects that attack trees.
Pisonia tree forest – Wikimedia
The tree most affected is pisonia – Pisonia grandis.
The principal insect is another scale, Pulvinaria urbicola. There are some reports that the scale is farmed by ants; species mentioned include several introduced species such as the yellow crazy ant, Paratrechina longicornis.
The scale is probably from the West Indies. Once it reached the Pacific, it might have been distributed to additional islands on seabirds, which travel long distances between the atolls.
The scale’s impact is unclear.
At first, in the mid-2000s, impacts seemed dire. It was reported to be causing widespread tree death on Palmyra and Rose atolls, islands around northeastern Australia, in the Seychelles, and possibly in Tonga.
However, in 2018, scientists reported that eradication of rats on Palmyra Atoll had resulted in an immediate spurt of reproduction of a tree. Numbers of “native, locally rare tree” seedlings (possibly but not explicitly said to be Pisonia grandis) jumped from 140 pre-eradication to 7,756 post-eradication (in 2016). The study made no mention of the scale.
Rose Atoll has only one small island (6.6 ha) with vegetation. Before 1970, it was dominated by Pisonia grandis, but by 2012, there were only seven trees on the island. Several possible causes of this decline have been suggested. Other than the scale, suggested causes include storms, drought, rising sea level / saltwater incursion, and imbalance of bird guano-derived nutrients in the soil. [All information about Rose Atoll is from Peck et al., 2014)
A survey carried out in April 2012 and November 2013 detected 73 species of arthropods from 20 orders on Rose Island, including nine ant species (all but one non-native). Two of these ants – Tetramorium bicarinatum and T. simillimum – were detected tending the scales on Pisonia.
The survey found no evidence of natural enemies of the Pulvinaria scales.
The scientists tested treatment of Pisonia with the systemic insecticide imidacloprid. This treatment apparently reduced scale populations considerably for several months, but then they began to build up again.
In contrast to Palmyra, Polynesian rats (Rattus exulans) were eliminated from Rose Atoll in 1990–1991 – so their role in destroying the trees had ended 20 years before the study. What does the continued decline of the Pisonia trees in subsequent decades suggest for the future of Pisonia trees on Palmyra?
I have sought updates on the tree-pest situations on Guam and the other Pacific islands, but my queries have not received a reply.
CABI November 2019. Aulacaspis yasumatsui (cycad aulacaspis scale (CAS)) or the Asian cycad scale. https://www.cabi.org/isc/datasheet/18756 (was formerly Commonwealth Agricultural Bureaux (CAB) International; now apparently just uses acronym)
Marler, T.E. pers. comm. August 15, 2012
Marler, T.E. 2010. Cycad mutualist offers more than pollen transport. American Journal of Botany, 2010; 97 (5): 841. Viewed as materials provided by University of Guam, via EurekAlert; accessed 6 August, 2012.
Marler, T., Haynes, J. & Lindstrom, A. 2010. Cycas micronesica. The IUCN Red List of Threatened Species 2010: e.T61316A12462113. http://dx.doi.org/10.2305/IUCN.UK.2010-3.RLTS.T61316A12462113.en Accessed 22 April, 2020.
Marler, T.E., and A. Moore. 2010. Cryptic Scale Infestations on Cycas revoluta Facilitate Scale Invasions. HortScience. 2010; 45 837-839. Retrieved August 6, 2012 from www.eurekalert.org
Marler, T.E. and K.J. Niklas. 2011. Reproductive Effort and Success of Cycas micronesica K.D. Hill Are Affected by Habitat. International Journal of Plant Sciences, 2011; 172 (5): 700. Viewed as materials provided by University of Guam, via EurekAlert; accessed 6 August, 2012.
Peck, R., P. Banko, F. Pendleton, M. Schmaedick, and K. Ernsberger. 2014. Arthropods of Rose Atoll with Special Reference to Ants and Pulvinaria urbicola scales (Hemiptera: Coccidae) on Pisonia grandis trees. Hawaii Cooperative Studies Unit. University of Hawaii. Technical Report HCSU-057 December 2014
vanWilgen, B.W.,J. Measey, D.M. Richardson, J.R. Wilson, T.A. Zengeya. Editors. 2020. Bioinvasions in South Africa. Invading Nature. Springer Series in Invasion Ecology 14.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm These reports do not include details on the pest situation on the Pacific islands (including Hawai`i).
EAB-kiled ash tree in Shenandoah National Park in 2016 photo by F.T. Campbell
As we have known for years, forests of the eastern United States are under severe pressure from non-native forest insects and diseases. Several recent studies have put this fact into perspective.
Fei et al. (2019) found that the 15 most damaging introduced species threaten 41.1% of the total live forest biomass in the 48 conterminous states. Nine of the 15 species included in this calculation are pests of the eastern forest. Indeed, the greatest increase in biomass loss, as measured by USDA Forest Service Forest Inventory and Analysis (FIA) plot data occurred here. Compensatory growth in unaffected trees and the recruitment of new regeneration occurs only later – as much as two or more decades after the pest invasions began. Fei et al. (2019) expect these losses will be exacerbated in the future due in part to the likelihood that additional pests will be introduced.
Randall Morin found that non-native pests had caused approximately 5% increase in total mortality, by tree volume, nation-wide.
Most widespread pest threats in the East
Scientists have used several methods of measuring introduced pests’ impacts. One measure is the number of counties where the pest is present. A second measure is the proportion of the volume of the host that has been affected. Both metrics are used by Morin. A third method, used by the CAPTURE Project (Potter et al. 2019a), is the number of hosts affected by the pest.
Morin and colleagues found that the European gypsy moth has invaded 630 counties – or 29% of the volume of its principal host, oaks. (In both cases, the gypsy moth trailed white pine blister rust in extent of infestation. The latter is nationwide but having its greatest impacts in the West). The CAPTURE Project found that the gypsy moth affected the largest number of hosts – 65.
Using the “counties invaded” metric, Morin and colleagues found that dogwood anthracnose had invaded 609 counties in the East (and additional areas in the West); the emerald ash borer had invaded 479 counties at the time of analysis; the hemlock woolly adelged had invaded 432 counties. Using the number of hosts impacted measure, oak wilt (Bretziella fagacearum) affected the second largest number of hosts – 61 (Potter et al. 2019a). [All these pests are described briefly here.]
Project CAPTURE (Potter and colleagues 2019a) evaluated 339 serious pests threatening one or more of 419 native tree species in the continental US. They included both native and introduced pests. They analyzed 1,378 pest-host combinations. They found that:
54% of the host tree species (228) are infested by an exotic pest – although only 28% of the 1,378 host/agent combinations involved pests are known to be non-native in origin.
Exotic agents have, on average, considerably more severe impacts than native pests.
Non-native pests had greater average severity on angiosperms than on conifers. (As an earlier blog documented, Mech and colleagues have reached a similar – although tentative – conclusion.)
Their estimate of the threat posed by non-native pests to forests – especially for the East – is an underestimate because established pests could spread to additional vulnerable areas and there is a high likelihood that new pests will be introduced. The Southeast was consistently a “coldspot” – despite the near extirpation of one understory tree – redbay.
Potter et al. (2019a) ranked forest threats in two ways. Four host families were at highest risk to alien pests, as measured by both the numbers of tree species affected and by the most host/agent combinations: Fagaceae (oaks, tanoaks, chestnuts, beech); Pinaceae (pines); Sapindaceae (soapberry family; includes maples and buckeye); Salicaceae (willows, poplars, aspens). When host families were ranked by the severity of the host/pest threat, Fagaceae was still at greatest risk, and Sapindaceae was still in the top four; however, Ulmaceae (elms) and Oleaceae (includes Fraxinus) replaced pines and willows.
A very interesting study was published by scientists based in the Blue Ridge Mountains of Virginia (Anderson-Teixeira et al. 2020). They contend that their area is a good example of what is happening more broadly in the Mid-Atlantic region.
Anderson-Teixeira et al. (2020) found that non-native pests have substantially impacted at least 24% of the 33 tree genera (eight genera) recorded as present in their study plots. They estimated that over the century beginning with the appearance of chestnut blight in the region and ending with the expected extirpation of ash trees, net live aboveground biomass (AGB) loss among affected species totaled roughly 6.6–10 kg m -2. Forty to sixty percent of this loss started before the Park initiated quantitative surveys of permanent plots in 1987. The authors estimated that chestnut contributed up to 50% of estimated AGB losses over the century. Consequently, the estimate has very high uncertainty.
Despite these losses, Anderson-Teixeira et al. (2020) found that both total aboveground biomass and diversity within individual study plots had largely recovered through increases in non-vulnerable genera.
Average above ground biomass across the plots established in Shenandoah National Park increased as the forest recovers from logging, farming, and other disturbances before formation of the Park. These increases were due primarily to reproduction and growth of tulip poplar (Liriodendron tulipifera) and growth (but not reproduction) of oaks. Net AGB biomass was lost in oak- and hemlock-dominated plots. At plots established in the neighboring Smithsonian Conservation Biology Institute, pests had caused relatively minor impacts on AGB.
Diversity of tree species also did not change much. In the Park, the average number of genera per plot declined only 3% between 1991 and 2013. Diversity at the landscape scale increased by two genera – from 26 to 28. Many individual plots, though, lost three genera due to non-native pests – chestnut, redbud, and hemlock. A fourth genus was lost due to stochastic change. At the same time, the plots gained six native genera). This finding might be skewed by the short duration of the study period, which missed initial declines in several taxa and captured only the initial stages of decline in ash.
Several taxa were lost from the monitoring plots but were not completely extirpated from the region. Even those species not “lost” suffered elevated mortality rates and steep declines in abundance and above-ground biomass. These declines have not been reversed. The exception was some oaks, which regained above ground biomass, but not abundance, following the gypsy moth outbreak in the 1980s and early 1990s.
Fei et al. (2019) found that losses in biomass due to non-native pests – as measured by FIA plot data – was greatest for ashes, elms, beech trees, and hemlocks..
Morin and colleagues found annual mortality rates had increased three-fold above background levels for ash, beech, and hemlock. They also calculated the present mortality rates for several species for which the majority of loss occurred before their study (consequently, they could not calculate a pre-invasion “background” rate to which present rates could be compared). These included American chestnut (mortality rate of 7%), butternut (mortality rate of 5.6%), and elm trees (mortality rate of 3.5%).
The CAPTURE Project (Potter et al. 2019a) identified fifteen host-agent combinations with the highest severity. Ten of these species are found in the Mid-Atlantic region:
American chestnut (Castanea dentata)
Allegheny chinquapin (C. pumila)
Carolina ash (Fraxinus caroliniana) ,
pumpkin ash (F. profunda)
Carolina hemlock (Tsuga caroliniana)
butternut (Juglans cinerea)
eastern hemlock (Tsuga canadensis)
white ash (Fraxinus americana)
black ash (F. nigra)
green ash (F. pennsylvanica)
Four of these species are in genera included among the eight genera evaluated in the study conducted in the Blue Ridge (Anderson-Teixeira et al. 2020): American chestnut, butternut, eastern hemlock, green and white ash. The four other genera in the Blue Ridge study were elm (Ulmus), oak (Quercus), redbud Cercis, and dogwood (Cornus). All except redbud are recognized by other sources as heavily affected by non-native pests – confirming Anderson-Teixeira et al. (2020)’s conclusion that findings on the Blue Ridge reflect the wider situation.
Anderson-Teixeira et al. (2020) note that several of these tree species have been declared imperiled by the International Conservation Union (IUCN): American chestnut, butternut, American elm, eastern hemlock, and ash species.
Anderson-Teixeira et al. (2020) report data on three taxa previously important in the canopy of Blue Ridge forests – chestnut, elms, and butternut. Chestnuts larger than 10 cm DBH had disappeared from the future site of Shenandoah National Park by 1910. Short-lived sprouts continue to be present in plots in the low-elevation Smithsonian Conservation Biology Institute. Two elm species were described as ‘‘sparse’’ in the 1939 qualitative survey. Elms have persisted at low densities, low biomass, and increasingly small sizes. Butternut was ‘‘common’’ in 1939, but had disappeared from Shenandoah NP by 1987. On the Smithsonian’s property, butternut declined from four living individuals in 2008 to two in 2018. The near disappearance of butternut reflects the national picture: FIA data show the species has decreased about 58% across its U.S. range since the 1980s – which is decades after butternut canker started having a detectable impact in the Midwest.
In the Park, oak-dominated plots lost on average 24.9% of individuals and 15% of aboveground biomass. After 1995, when the gypsy moth was better controlled by spraying of Bacillus thuringiensis var. curstaki, oak aboveground biomass increased gradually, driven by individual tree growth rather than new recruitment. Continued declines in oak abundance are attributable to oak decline and management actions (or inactions) that do not promote regeneration.
In a separate study, a group of oak experts went through a process of queries to identify the greatest threat to oaks now and in the future (Conrad et al. 2020). They initially identified the following threats as most important currently (descending order): gypsy moth, oak wilt, oak decline, climate change, and drought. The top five future threats were initially identified as climate change, oak wilt, sudden oak death, oak decline, and some unknown new or emerging (non-native) pest or pathogen. By the third round, after the experts thought about their colleagues’ responses, oak decline had replaced gypsy moth as the most critical threat currently. Attack by an unknown new or emerging (non-native) pest or pathogen replaced climate change as the most critical future threat. While there was not a complete consensus, the consensus was stronger on the threat from a new pest.
remnant eastern hemlock at Linderlost, Shenandoah National Park photo by F.T. Campbell
Anderson-Teixeira et al. (2020) reported that eastern hemlock was initially present in ten of Shenandoah plots, but was no longer recorded in the survey plots after 2007. (More than 20,000 insecticide-treated trees remain alive throughout Shenandoah NP).
Before arrival of the emerald ash borer, ash aboveground biomass was increasing in Shenandoah NP and stable on the Smithsonian Institute. EAB-caused mortality was first detected at the Smithsonian site in 2016 and accelerated steeply thereafter, exceeding 12.5% year by 2018. As of 2019, ash had lost 28% of individuals and 30% of aboveground biomass relative to 2016. Ninety-five percent of remaining live trees were considered “unhealthy’’ (Anderson-Teixeira et al. 2020).
eastern (flowering) dogwood; photo by F.T. Campbell
Unlike many studies, the Shenandoah study included understory species. Flowering dogwood declined by up to 90% from plots on the Smithsonian property; 2008–2019 mortality rates averaged 7.1%. Redbud declined by up to 76% from 1995 to 2018. The 2008–2019 mortality rates averaged 6.2% year.
Anderson-Teixeira et al. (2020) concede difficulty in estimating mortality due to less virulent or lethal pathogens, including Neofusicoccum spp. on redbud and Dutch elm disease on slippery elm.
Nevertheless, they believe their analysis probably underestimates the overall pest impacts because they did not analyze several other pest/host combinations known to be present in the Park: balsam woolly adelgid (Adelges piceae) on high-elevation populations of Abies balsamea; white pine blister rust (Cronartium ribicola) on eastern white pine (Pinus strobus); beech bark disease (Neonectria spp.) on American beech (Fagus grandifolia); thousand canker disease on walnut and butternut; and emerald ash borer on the novel host fringetree Chionanthus virginicus.
Another possible threat to oaks, winter moth (Operophtera brumata), is apparently now being controlled by the biocontrol agent Cyzenis albicans.
I am uncertain about the current status of two Diplodia fungi – Diplodia corticola and D. quercivora – link to blog which have been detected in both Florida and California. In Florida, almost all the symptomatic trees grow in cultivated settings where they are exposed to various stresses (Mullerin and Smith 2015).
However, host range studies indicate that 33 species of oaks and one species of chestnut that grow in the Southeast are vulnerable, to varying degrees, to D. corticola. Oaks in the red oak group (Section Lobatae) are more vulnerable than are white oaks (Section Quercus) (Mullerin and Smith 2015). In the test, the most vulnerable appear to be the following species native to the Southeast: Q. laurifolia, Q. virginiana, Q. geminata, Q. chapmanni, Q. laevis (turkey oak), Q. phellos, Q. pumila, and Q. incana (Dreaden et al. 2016).
What should we do?
Fei et al. (2019) noted that the losses to biomass would be exacerbated by the likely introduction of additional pests. They did not recommend any prevention actions.
Conrad et al. (2020) said their findings “lend support to national regulatory and awareness efforts to prevent the introduction and establishment of novel exotic insects and pathogens.”
Anderson-Teixeira et al. (2020) join others in declaring that future survival of the IUCN-listed species probably depends on conservation and restoration actions. They cite several sources, but not the CAPTURE Project – although the two studies reinforce each other. They specifically mention limiting invasive species’ spread through strengthened regulations and “enhanced plant biosecurity cyberinfrastructure”.
This last recommendation reinforces the message of Bonello et al. (2019) link to publication. We called for creation of a federal Center for Forest Pest Control and Prevention to implement end-to-end responses to forest pest invasions. One focus would be correcting the currently-inadequate focus on detection, development and deployment of genetic resistance while using modern techniques that allow for much faster breeding cycles.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
SOURCES
Anderson-Teixeira, K.J., V. Herrmann, W.B. Cass, A.B. Williams, S.J. Paull, E.B. Gonzalez-Akre, R. Helcoski, A.J. Tepley, N.A. Bourg, C.T. Cosma, A.E. Ferson, C. Kittle, V. Meakem, I.R. McGregor, M. N. Prestipino, M.K. Scott, A.R. Terrell, A. Alonso, F. Dallmeier, and W.J. McShea. Date? Long-Term Impacts of Invasive Insects and Pathogens on Composition, Biomass, and Diversity of Forests in Virginia’s Blue Ridge Mountains. Ecosystems
Bonello, P. , F.T. Campbell, D. Cipollini, A.O. Conrad, C. Farinas, K.J.K. Gandhi, F.P. Hain, D. Parry, D.N. Showalter, C. Villari, and K.F. Wallin. 2019. Invasive tree pests devastate ecosystems – A proposed new response framework. Frontiers
Conrad, A.O., E.V. Crocker, X. Li, W.R. Thomas, T.O. Ochuodho, T.P. Holmes, and C. D. Nelson. 2020. Threats to Oaks in the Eastern US: Perceptions and Expectations of Experts. Journal of Forestry, 2020, 14–27
Dreaden, Black, Mullerin, and Smith. Poster presented at the 2016 USDA Invasive Species Research Forum
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. Liebhold. 2019. Biomass losses resulting from insect and disease invasions in United States forests. Proceedings of the National academy of Sciences.
Guo, Q., S. Feib, K.M. Potter, A.M. Liebhold, and J. Wenf. 2019. Tree diversity regulates forest pest invasion. PNAS. www.pnas.org/cgi/doi/10.1073/pnas.1821039116
Morin, R.S., K.W. Gottschalk, M.E. Ostry, A.M. Liebhold. 2018. Regional patterns of declining butternut (Juglans cinerea L.) suggest site characteristics for restoration. Ecology and Evolution.2018;8:546-559
Morin, R. A. Liebhold, S. Pugh, and S. Fie. 2019. Current Status of Hosts and Future Risk of EAB Across the Range of Ash: Online Tools for Broad-Scale Impact Assessment. Presentation at the 81st Northeastern Forest Pest Council, West Chester, PA, March 14, 2019
Mullerin, S. & J.A. Smith. 2015. Bot Canker of Oak in FL Caused by Diplodia corticola & D. quercivora. Emergent Pathogens on Oak and Grapevine in North America. FOR318
Potter, K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019a. Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests. 2019 10 304.
Potter, K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019b. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation. (2019)
Dream Lake, Rocky Mountain National Park, with limber pine photo by F.T. Campbell
In this blog, I summarize two pest threats to the unique ecosystems on high-elevation mountain ridges in the West. At risk are several keystone tree species: the five-needle pines growing at high elevations (“high-five” pines) and subalpine fir. The invasive species causing this damage – white pine blister rust (WPBR; Cronartiumribicola) and balsam woolly adelgid (BWA; Adelges piceae) – are two of the most widespread non-native species threatening North American trees and affecting the highest proportion of host volumes (Morin).
The pines being killed by white pine blister rust are whitebark pine (Pinus albicaulis), limber pine (P. flexilis), Rocky Mountain bristlecone pine (P. aristata), foxtail pine (P. balfouriana), and southwestern white pine (P. flexilis var. reflexa). As of 2010, infestations had not been reported on Great Basin bristlecone pine (P. longaeva) and the Mexican white pine species. [Unless otherwise indicated, information on white pine blister rust is from a comprehensive review and synthesis published in the August 2010 issue of Forest Pathology (Vol. 40:3-4).]
As noted above, sub-alpine fir (Abies lasiocarpa) is also being affected – although less uniformly than the pines – by the balsam woolly adelgid.
Both of these pests arrived approximately a century ago, but they are still spreading and causing additional damage. White pine blister rust had spread widely throughout the West within 40 years of its introduction. Meanwhile, BWA spread among lowland and subalpine firs along the Pacific coast from California to British Columbia within 30 years of its first detection. Its spread eastward was slower, but relentless. It reached Idaho, Montana, Utah and interior British Columbia within 50 years. Also, BWA reached Alaska within 90 years of its introduction in California. These pests are perfect examples of how invasive species introduced long ago are dreaded “gifts that keep on giving”.
For a detailed discussion of these pests’ impacts, see the descriptions posted here. To summarize, though, WPBR is present in the ranges of eight of the nine vulnerable western white pines and has caused severe mortality to some species (Sniezko et. al. 2011). For example, 88% of the limber pine range in Alberta is affected (Dawe et al. 2020). WPBR is generally causing more damage to its hosts’ northern populations. Impact of the BWA are more subtle than WPBR. Also, impacts’ severity is linked to climatic conditions. For example, measurable decline on the Olympic Peninsula was greater on south-facing slopes. However, the study did not determine whether this reflected heat-loading and tree stress or more abundant subalpine fir on these slopes. An estimated 19-53% (average 37%) of subalpine fir trees had died on sample plots on one ridge over the 19 years since BWA was first detected there. Overall forest growth after 2007 could indicate partial recovery, a momentary pause in BWA invasion, or tree growth after severe weather events (Hutton 2015).
Ranges of Trees at Risk
Many of the host trees of these two pests are widespread; others are more narrowly endemic.
Limber pine reaches from Alberta and British Columbia south to mountain peaks in Arizona and New Mexico. Whitebark pine is found from Alberta and British Columbia to California and Nevada (USDA Plants database. Subalpine fir stretches from southeast Alaska along the Canadian Rockies coast into Washington, Oregon, east into Idaho, Montana, Wyoming, Colorado, Utah, even into scattered mountain ranges of Nevada and New Mexico (Hutton 2015).
Limber pine and subalpine fir are also found in a wide range of ecosystems within these ranges. Limber pine is found at both upper and lower tree lines in grassy, open forests; on exposed rocky slopes; and in dense, mixed-conifer stands. Subalpine fir is a pioneer species on ridges, alpine meadows, avalanche chutes, and lava beds (Ragenovich and Mitchell, 2006).
Before arrival of non-native pests or pathogens, these tree species have persisted for thousands of years under harsh conditions (Hutton 2015). Many of the individual trees were long-lived; some five-needle pines, e.g., bristlecone pines, have famously live for thousands of years. Core studies demonstrated that subalpine firs trees could live 272 years in the forests of Olympic National Park and 240 years in Glacier National Park (Hutton 2015). Surely loss of these trees – or even their conversion from large and old to small and short-lived – will result in significant destruction of these unique biomes.
All these trees play important roles in high altitude, unique ecosystems (Pederson et al. no date; Dawe 2020; Hutton 2015):
They retain ground water, slow the rate of snow melt, and maintain stream flow characteristics and water quality;
They curtail soil erosion and maintain slope stability; and
They provide high-value food and shelter to wildlife.
Whitebark and limber pines are famous for providing critical food for many wildlife species at high elevations —notably bears and nutcrackers (Compendium and Dawe 2020).
More Pest Threats
Other diseases, insects, and disturbances also pose serious threats to these tree species. The threats vary by region and age of the stand. They include – for the pines — mountain pine beetle (Dendroctonus ponderosae), dwarf mistletoe (Arceuthobium spp.), and various shoot, cone or foliage insects and pathogens. For subalpine fir, threats include western balsam bark beetle (Dryocoetes confusus), fir engraver (Scolytus ventralis), and the fir root bark beetle (Pseudohylesinus granulatus) (Hutton 2015). Trees are also damaged by bear and deer, seed predation by squirrels, wildfire, and biotic succession.
On Washington’s Olympic Peninsula, BWA initiates or predisposes subalpine fir for a novel disturbance complex. BWA-caused stress makes the trees more susceptible to moisture stress and endemic bark beetle attack. Surviving trees are subsequently subject to toppling by wind. A tree can die in a few years, survive with insects for up to 20 years, or recover, depending on duration, severity, and location of infestation, and local environmental conditions (Hutton 2015).
BWA study plots in the Cascade Range experienced subalpine fir mortality ranging from 7 to 79% (measured as stem counts, not basal area) over a 19 to 38 years study period. Higher mortality occurred at low-elevation, mesic sites. One stand experienced 40% mortality in 19 years, but lost the remaining 60% during a subsequent spruce budworm infestation. Most plots continued to show sporadic signs of adelgid presence and continued tree mortality. However, 41-69% of trees survived stem infestations (Hutton 2015).
How to Protect These Ecosystems
The seeds of both whitebark and limber pines are dispersed to newly disturbed, open areas by Clark’s nutcracker (Nucifraga columbiana). Furthermore, whitebark cones open to release seeds only after fire. This had led to expectations that prescribed fire could promote regeneration of these species. However, studies by Dawe (2020) and other have found that nutcracker seed caching behavior and seedling establishment are complex. Fire management might have to vary among regions, demanding consideration of stand characteristics,like openness and the presence of other tree species. For example, in the Colorado Front Range, limber pine can be replaced by subalpine fir when fire-free intervals are long. On the other hand, in Alberta, fire appeared to boost regeneration of the dominant tree species in the stands pre-fire. In the study areas, these were white spruce (Picea glauca) and lodgepole pine (Pinus contorta) (Dawe 2020). Dawe recommends protecting existing stands of limber pine through fire mitigation efforts, e.g., thinning and other fuel treatments, and supplementary planting of seedlings.
Efforts to find biocontrol agents to target the balsam woolly adelgid began in 1957; the original focus was on the insects’ damage to Fraser fir (Abies fraseri) in the southern Appalachians. More than 25 predatory species have been introduced from Europe and Asia. There was simultaneous research on native predators. None has had an impact on BWA populations in either the East or the West.
Neither white pine blister rust nor balsam woolly adelgid is considered a quarantine pest by federal officials, so there is no attempt to prevent their movement via interstate trade in Christmas trees, timber, or nursery stock. Hutton (2015) hypothesizes that the absence of regulatory measures targetting BWA arises from the pest’s gradual effect and the hosts’ not being commercially important as timber species (although several firs are important in horticulture and as Christmas trees). I think another factor is that the pests were introduced so long ago and are now widespread.
Efforts are under way to detect resistant genotypes to be used in breeding programs. Several of the lower-elevation five-needle pines vulnerable to WPBR have benefitted from extensive breeding efforts Whitebark pine has more recently been added to programs.
The eastern Fraser fir is the target of breeding – primarily for Christmas trees (APS). However, at least small-scale volunteer efforts have been carried forward by the Alliance for Saving Threatened Forests.
Hutton (2015) expresses hope that evolutionary pressure by BWA might enhance survival of more resistant forms of subalpine fir and lead to their gradual takeover. However, I ask, why leave it to chance?
In this context, I remind you of my involvement with a group (see Bonello et al. 2019) proposing creation of a federal Center for Forest Pest Control and Prevention to implement end-to-end responses to forest pest invasions – including overcoming the currently inadequate focus on detection, development and deployment of genetic resistance using modern techniques that allow for much faster breeding cycles.
I am puzzled that the Project CAPTURE places whitebark pine and subalpine fir only in Class A4.2, not among the highest priority species (Potter et al. 2019). As I blogged last spring, Project CAPTURE is part of a multi-partner effort to categorize and prioritize US tree species for conservation actions based on the threats and the trees’ ability to adapt to those threats. I find it puzzling because I am not sure I agree that these two species have a moderately high mean pest severity score – as required by the category. I am less puzzled by the assignment of a low adaptive capacity score.
Limber pine apparently ranks even lower in the Project CAPTURE priority process.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
SOURCES
A comprehensive review and synthesis of the history, ecology, and management of white pines threatened by white pine blister rust see the August 2010 issue of Forest Pathology (Vol. 40:3-4).
Bonello, P. , F.T. Campbell, D. Cipollini, A.O. Conrad, C. Farinas, K.J.K. Gandhi, F.P. Hain, D. Parry, D.N. Showalter, C. Villari, and K.F. Wallin. 2019. Invasive tree pests devastate ecosystems – A proposed new response framework. Frontiers
Dawe, D.A., V.S. Peters, M.D. Flannigan. 2020. Post-fire regeneration of endangered limber pine (Pinus flexilis) at the Northern extent of its range. Forest Ecology and Management 457 (2020) 117725
Hutton, K.M. 2015. A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy. University of Washington. Available here
Morin, R. Presentation to the 81st Northeastern Forest Pest Council Northeastern states forst agencies, Philadelphia, Pennsylvania, March 2019.
Potter, K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S. 2019. Prioritizing the conservation needs of US tree spp: Evaluating vulnerability to forest P&P threats, Global Ecology and Conservation (2019), doi: https://doi.org/10.1016/
Sniezko, R.A., M.F. Mahalovich, A.W. Schoettle, D.R. Vogler. 2011. Past and Current Investigations of the Genetic Resistance to Cronartium ribicola in High-elevation Five-needle Pines. In Keane, R.F., D.F. Tomback, M.P. Murray, and C.M Smith, eds. 2011. The future of high-elevation, five-needle white pines in Western North America. Proceedings of the High Five Symposium. 28-30 June, 2010. Missoula, MT.
The California Department of Food and Agriculture (CDFA) is seeking comments on the appropriate pest rating for Leptosillia pistaciae, a recently discovered fungus that causes pistachio canker.
The
Department’s draft pest ranking assigns the highest Economic Impact score –
three. It assigns a medium Environmental Impact – two. This is because the
pathogen can kill an important native shrub, with possible follow-on
consequences of reduced biodiversity, disrupted natural communities, or changed
ecosystem processes.
CDFA
states that there is no uncertainty in its evaluation, but I see, and describe
here, numerous questions about the possible true extent of the invasion and
possible host range.
Comments are due
on April 4, 2020.
The
pathogen was detected in June 2019, when a habitat manager from an ecological
reserve in San Diego County noticed multiple dead lemonade berry shrubs (Rhus integrifolia) in one of the parks.
This is the first known detection of Leptosillia
pistaciae in the United States and on this host. USDA APHIS has classified Leptosillia pistaciae as a federal quarantine
pest. Rhus and Pistacia are in the same family, Anacardiaceae (cashews and sumacs).
According
to the CDFA, Leptosillia pistaciae is
the only member of this fungal genus known to be associated with disease
symptoms on plants. Other species are endophytes or found in dead plant
tissues. [It is not at all unusual for fungal species to be endophytes on some
plant hosts but pathogenic on others. A California example is Gibberella
circinata (anamorph Fusarium circinatum), which causes
pitch canker on Monterey pine (Pinus
radiata) but is an endophyte on various grass species (Holcus lanatus and Festuca
arundinacea).]
(Reminder: this is the second new pest of native species detected in California state in 2019; I blogged about an ambrosia beetle in Napa County here. )
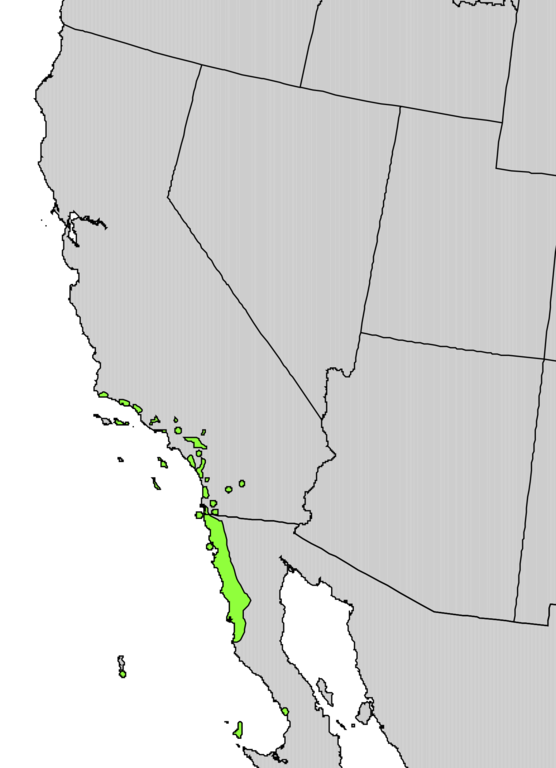
Rhus integrifolia (lemonade berry
or lemonade sumac) is native to California. It grows primarily in the south, along
the coast – from San Diego to San Luis Obispo. However, some populations are
also found in the San Francisco Bay area. This and other sumacs are also sold
in the nursery trade.
On
pistachio trees in Italy, symptoms are observed in the winter and late spring. During
the winter dormant season, trees had gum exudation and cracking and peeling of
bark on trunks and branches. On trunks and large branches, cankers appeared
first as light, dead circular areas in the bark; subsequently they became
darker and sunken. Under the bark, cankers were discolored with necrotic
tissues; in some cases, these extended to the vascular tissues and pith. During
the active growing season, the symptomatic plants also showed canopy decline.
Inflorescences and shoots, originating from infected branches or twigs, wilted
and died. When the trunk was girdled by a canker, a collapse of the entire tree
occurred.
range map for Rhus integrifolia
On
lemonade berry, large clumps of dead
adult shrubs were observed on the edge of hiking trails. Some shrubs that had completely
dead foliage were re-sprouting from their bases. Trunks of shrubs that were not
completely dead were copiously weeping sap and fluids and showed foliage
browning and die back with symptoms of stress.
It
is thought that spores could be spread by wind, rain splashing, and the
movement of dead or dying trees, greenwaste, and infected nursery stock. Contaminated
pruning tools might also transport the spores. The possibility of a latent
phase – or perhaps asymptomatic hosts – adds to the probability of
anthropomorphically assisted spread.
I question how much effort has been put into detection surveys, especially in natural systems with native Rhus species. California has three other native sumacs: R. ovata, R. aromatica, and Malosma laurina (CNPS; full citation at the end of the blog). In addition, there are numerous other species in the family, including poison oaks (Toxicodendron spp.) and the widespread invasive plant genus Schinus.
Furthermore, some plants in the family (other than pistachios) are grown for fruit or in ornamental horticulture, including two of the native sumacs and two non-native species, Rhus glabra and R. lanceolata, cashew, mango, and smoke trees (Cotinus spp.).
Yet
CDFA confidently states that there are only two hosts and that it has been
detected in only one population – that in San Diego. This is because CDFA
considers only official records identified by a taxonomic expert and supported
by voucher specimens.
CDFA
states that the pathogen is likely to survive in all parts of the state where
pistachios are grown – primarily in the Central Valley. California supplies 98%
of the pistachios grown in the United States; the remainder is raised in
Arizona and New Mexico. California production occurred on 178,000 acres in
2012. A map is included in a flyer on production available at the url listed at
the end of this blog.
In
discussing spread potential, no mention is made of possible human-assisted
spread.
The CDFA document includes instructions for submitting comments; the deadline is April 4.
Sources:
Rhus and related
species native to California: California Native Plant Society
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
prickly pear cacti in Big Bend National Park photo by Blake Trester, National Park Service
The cacti that are such important components of desert ecosystems across nearly 2 million square miles straddling the U.S.-Mexico border are under threat from non-native insects – as I have noted in earlier blogs. Of course, cacti are important in other ecoregions, too – I wrote recently about the columnar cacti in the dry forests of Puerto Rico.
Flat-padded prickly pear cacti of the genus Opuntia are threatened by the cactus moth, Cactoblastis cactorum.
In 1989, the cactus moth was found
in southern Florida, to which it had spread from the Caribbean islands (Simonson
2005). Recently, the moth was found to have spread west as far as the Galveston,
Texas, area and near I-10 in Columbus, Texas, about 75 miles west of central
Houston (Stephen Hight, pers. com.) Two
small outbreaks on islands off Mexico’s Caribbean coast have been eradicated.
In Florida, the cactus moth has
caused considerable harm to six native species of prickly pear, three of which
are listed by the state as threatened or endangered.
When the cactus moth reaches the
more arid regions of Texas, it is likely to spread throughout the desert
Southwest and into Mexico. In the American southwest, 31 Opuntia species are at risk; nine of them are endemic, one is endangered.
Mexico is the center of endemism for the Opuntia
genus. In Mexico, 54 Opuntia species
are at risk, 38 of which are endemic (Varone et al. 2019; full citation at end of this blog).
The
long-term effects of the cactus moth on these North American Opuntia are unknown because there may be
substantial variations in tolerance. The attacks observed in the Caribbean
islands have shown great variability in various cactus species’ vulnerability (Varone et al. 2019).
The Opuntia cacti
support a diversity of pollinators as well as deer, javalina (peccaries),
tortoises, and lizards. Prickly pears also shelter packrats and nesting birds (which
in turn are fed on by raptors, coyotes, and snakes), and plant seedlings. Their
roots hold highly erodible soils in place (Simonson 2005).
While scientists have been concerned about the possible impacts of the cactus moth since it was detected in Florida 30 years ago, a substantial response began only 15 years later. The U.S. Department of Agriculture began trying to slow the spread of the cactus moth in 2005 (Mengoni Goñalons et al. 2014), with a focus on surveys and monitoring, host (cactus) removal, and release of sterile males. This program was successful at slowing the moth’s spread and eradicating small outbreaks on offshore islands of Alabama, Mississippi, and Mexico.
Cactus moth damage to native cacti in Florida photo by Christine Miller, UF/IFAS
However,
the moth continued to spread west and the program never received an appropriation from Congress. The primary funding source was a US – Mexico
Bi-National Invasive Cactus Moth Abatement Program. Both countries contributed
funds to support the research and operational program to slow the spread in the
U.S. Funds were provided through USDA Animal and Plant Health and Inspection
Service (APHIS) and the Mexican Secretariat of Agriculture, Livestock, Rural
Development, Fisheries and Food (SEGARPA). Unfortunately,
funding was reduced by both entities and became inadequate to maintain the
Bi-National Program.
Therefore, in 2012, APHIS abandoned
its regional program and shifted the focus to biocontrol. This is now
considered the only viable control measure in the desert Southwest where vulnerable
cacti are numerous and grow close together. The biocontrol project has been funded
since 2012 through the Plant Pest and Disease Management and Disaster
Prevention program (which receives funding through the Farm Bill). It has
received a total of slightly more than $2
million over seven years. More than half the funds went to the quarantine
facility to support efforts to rear non-target hosts and verify the biocontrol
agent’s host specificity. About a quarter of the funds supported complementary
work of an Argentine team (both the cactus moth and the most promising
biocontrol agent are native to Argentina). Much smaller amounts have supported
U.S.-based scientists who have studied other aspects of the cactus moth’s
behavior and collected and identified the U.S. moths being tested for their
possible vulnerability to attack by a biocontrol wasp.
Here are
details of what these dedicated scientists achieved in just the past seven
years at the relatively low cost of roughly $2 million. Unfortunately, the project now faces a funding crisis and
we need to ensure they have the resources to finish their work.
Some
Specifics of the BioControl Program
After literature reviews, extensive collections,
and studies in the cactus moth’s native habitat in Argentina (Varone et al. 2015), a newly described wasp, Apanteles
opuntiarum (Mengoni Goñalons et al. 2014), has been determined to be host
specific on Argentine Cactoblastis species and the most promising
candidate for biocontrol. Wasps were collected in Argentina and sent to
establish a colony in a quarantine facility in Florida to enable host
specificity studies on North American Lepidoptera (Varone et al. 2015).
Quarantine
host specificity studies and development of rearing technology has not been straightforward. Initially, it was
difficult to achieve a balanced male/female ratio in the laboratory-bred generations;
this balance is required to maintain stable quarantine laboratory colonies for
host range testing. This difficulty was overcome. A second challenge was high
mortality of the cactus-feeding insects collected in the Southwest that were to
be test for vulnerability to the biocontrol wasp. These desert-dwellers don’t
do well in the humid, air-conditioned climate of the quarantine facility! For
these difficult-to-rear native insects, scientists developed a molecular
genetics method to detect whether eggs or larvae of the cactus moth parasitoid were
present inside test caterpillars after they were exposed to the wasps. For easy
to rear test insects, caterpillars are exposed to the wasps and reared to
adulthood. Host specificity tests have been conducted on at least five species
of native U.S. cactus-feeding caterpillars and 11 species of non-cactus-feeding
caterpillars (Srivastava
et al. 2019; Hight pers.comm.).
To
date there has been no instance of
parasitism by Apanteles opuntiarum on either lepidopteran non-target species or
non-cactus-feeding insects in the Florida quarantine or in field collections in
Argentina (Srivastava et al.
2019; Varone et al. 2015; Hight pers.comm.).
The scientists expected to complete host-specificity testing in the coming months, then submit a petition to APHIS requesting the release of the wasp as a biocontrol agent. Unfortunately, the project’s request for about $250,000 in the current year was not funded. This money would have funded completion of the host specificity testing, preparation of a petition to APHIS in support of release of the biocontrol agent into the environment, and preparation of the release plan.
Meanwhile,
what can we expect regarding the probable efficacy of the anticipated biocontrol
program?
Some
of the wasp’s behavioral traits are encouraging. The wasp is widely present in
the range of the cactus moth, and persisted in these areas over the years of
the study. The wasp can deposit multiple eggs with each “sting”. Multiple wasps
can oviposit into each cactus moth without detriment to the wasp offspring. Unmated wasp females
produce male offspring only, whereas mated females produce mixed offspring
genders. In the field, female wasps attack cactus moth larvae in a variety of
scenarios: they wait at plant access holes to sting larvae when they come
outside to defecate; they attack larvae when they are moving on the surface of
the pads; they can sting the youngest cactus moth larvae through the thin plant
wall of mined the pads; and they enter large access holes created by older
larvae and attack larger larvae. The wasps are attracted by the frass
(excrement) left on the outside of the cactus pads by cactus moth larvae (Varone
et al. 2020).
However, I wonder about the extent
to which the cactus moth is controlled by parasitoids in Argentina. Cactoblastis eggs are killed
primarily by being dislodged during weather events (rain and wind) and by
predation by ants. First instar larvae are killed primarily by the native Argentine
cactus plants’ own defenses – thick cuticles and release of sticky mucilage when
the young larvae chew holes into the pads where they enter and feed internally.
As larvae feed and develop inside the pads, the primary cause of mortality is
natural enemies.
Of
all the parasitoid species that attack C. cactorum, A. opuntiarum
is the most abundant and important. When the larvae reach their final state (6th
instars), they leave the pads and find pupation sites in plant litter near the
base of the plants. It is at this stage that the parasitism from A.
opuntiarum is detected in the younger larvae that were attacked while
feeding inside pads. As the moth larva begins to spin silk into which to
pupate, larvae of the wasp erupt through the skin of the caterpillar and pupate
within the silk spun by the moth. Predation by generalists (ants, spiders,
predatory beetles) accounted for high mortality of the unprotected last instar
and pupae (Varone et al. 2019).
Finally,
the cactus moth has three generations per year when feeding on O. stricta in the subtropical and tropical coastal areas of the Americas
and the Caribbean. In Argentina, on its native host, the moth completes
only two generations per year (Varone et
al. 2019).
How to
Get the Program Support Needed
Opuntia in Big Bend National Park Photo by Cookie Ballou, National Park Service
To date, no organized
constituency has advocated for protection of our cacti from non-native insect pests.
Perhaps now that the Cactoblastis
moth is in Texas, the threat it represents to our desert ecosystems will become
real to conservationists and they will join the struggle. The first step is to
resolve the funding crisis so that the agencies can complete testing of the biocontrol
agent and gain approval for its release. So now there is “something people can
do” – and I hope they will step forward.
I hope Americans are not actually indifferent
to the threat that many cacti in our deserts will be killed by non-native
insects. Many are key components of the ecosystems within premier National
Parks, and other protected areas. Cacti also are beautiful treasures in
botanical gardens. I hope conservationists will agree that these threats must
be countered, and will help to ensure
funding of the final stages of the biocontrol tests.
Sources
Mengoni Goñalons, C., L. Varone, G. Logarzo, M. Guala, M.
Rodriguero, S.D. Hight, and J.E. Carpenter. 2014. Geographical range & lab
studies on Apanteles opuntiarum (hymenoptera: braconiDae) in AR, a candidate
for BC of Cactoblastis cactorum (Lepidoptera: Pyralidae) in North America. Florida
Entomologist 97(4) December 2014
Srivastava, M., P. Srivastava, R. Karan, A. Jeyaprakash,
L. Whilby, E. Rohrig, A.C. Howe, S.D. Hight,
and L. Varone. 2019. Molecular detection method developed to track the
koinobiont larval parasitoid Apanteles opuntiarum (Hymenoptera: Braconidae) imported from Argentina to control Cactoblastis cactorum (Lepidoptera:
Pyralidae). Florida Entomologist 102(2): 329-335.
Varone, L., C.M. Goñalons, A.C. Faltlhauser, M.E. Guala,
D. Wolaver, M. Srivastava, and S.D. Hight. 2020. Effect of rearing Cactoblastis cactorum on an artificial
diet on the behavior of Apanteles
opuntiarum. Applied Entomology DOI: 10.1111/jen.12731.
Varone,
L., G. Logarzo, J.J. Martínez, F. Navarro, J.E. Carpenter, and S.D. Hight.
2015. Field host range of Apanteles
opuntiarum (Hymenoptera: Braconidae) in Argentina, a potential biocontrol
agent of Cactoblastis cactorum
(Lepidoptera: Pyralidae) in North America. Florida Entomologist — Volume 98,
No. 2 803
Varone, L., M.B. Aguirre, E. Lobos, D. Ruiz Pérez, S.D. Hight, F. Palottini, M. Guala, G.A. Logarzo. 2019. Causes of mortality at different stages of Cactoblastis cactorum in the native range. BioControl (2019) 64:249–261
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
APHIS can protect our native & agricultural plants – but will it?
Imports
of large numbers of plants for planting from Southeast Asia represents a
significant biosecurity risk for forestry, horticulture, and natural ecosystems
in North America and Europe. This threat
is likely to grow unless APHIS takes
action under its emergency authorities.
Recent
pest introductions and related studies indicate that Southeast Asia is a newly-discovered
center of origin for plant pathogens. Places of particular concern are Vietnam,
southern Yunnan Province and Hainan Island of China, northern Laos, the eastern
Himalayas, and Taiwan. Significant pathogens and associated insects apparently centered
in these areas include the sudden oak death pathogen (Phytophthora ramorum) and other Phytophthora
species; and several ambrosia beetles and associated fungi, including the laurel
wilt fungus (Raffaelea lauricola) and
its primary vector (Xyleborus glabratus),
and the polyphagous (Euwallacea whitfordiaodendrus)
and Kuroshio shot hole borers (Euwallacea
kuroshio).
Southeast
Asia is attractive to the plant trade because of the region’s high floral diversity,
including such sought-after families as Ericaceae (rhododendrons). Indochina has
more than 10,350 vascular plant species in 2,256 genera – equaling more than
20% of the world’s plant species (Jung et
al. 2019).
Pathogens
are notoriously difficult to detect during inspections at the time of shipment.
One-time inspections of high volume imports are especially weak and prone to
failure.
How do we
protect America’s flora?
APHIS could — but has not yet — developed requirements that these countries institute integrated pest management procedures for their exporting nurseries – as provided under amendments to APHIS’ Q-37 regulation and ISPM#36. In any case, it is unlikely that such procedures would minimize the risk because many of the plants that would be imported would probably be wild-collected.
APHIS has – and should use – far more effective means to minimize risk. These are the Federal orders and listing process known as “not authorized for importation pending pest risk assessment” or NAPPRA. If – despite the scientific evidence – APHIS continues to allow high volumes of dangerous imports, the agency should immediately institute new phytosanitary controls to its inspection process. These include relying on risk-based inspection regimes and molecular high-through-put detection tools.
Supporting
Material
SOD-killed tanoaks in Big Sur; photo provided by Matteo Garbelotto, UC Berkeley
Phytophthora species
A
team of European pathologists, led by Thomas Jung and including Clive Brasier
and Joan Webber (see full citation at the end of this blog) surveyed Phytophthora species by sampling
rhizosphere soils in 25 natural and semi-natural forest stands, isolations from
naturally fallen leaves, and waters in 16 rivers in temperate and subtropical
montane and lowland regions of Vietnam during 2016 and 2017.
These
studies detected 13 described Phytophthora
species, five informally designated taxa, and 21 previously unknown taxa. Detections were made from soil samples
taken from 84% of the forest stands and from all rivers.
As I reported in am earlier blog, P. ramorum and P. cinnamomi were among those species detected. Both the A1 and A2 mating types of both P. ramorum and P. cinnamomi co-occurred.
The
survey also detected at least 15 species in other genera of oomycetes.
The
scientists conclude that most of the 35 forest Phytophthora species detected are native to Vietnam or nearby
surrounding areas, attributing species in Phytophthora
clades (taxonomically related groups) 2, 5, 6, 7, 8, 9, and 10 as native to
Indochina. Different clades were detected in high-elevation vs. lowland rivers, cooler (subtropical)
vs. tropical streams, and in soils vs. streams. Given the relatively
limited number and diversity of the sampled sites and ecosystem types, it is
likely that the true Phytophthora diversity
of Vietnam is markedly higher (Jung et al.
2019)
Worrying
diversity of Phytophthora has been
detected in other areas of Southeast Asia. A 2013 survey in natural forests and
streams of Taiwan detected 10 described species and 17 previously unknown taxa
of which 9 were of hybrid origin. In three areas in northern Yunnan, a Chinese
province adjacent to northern Vietnam, eight Phytophthora species were isolated from streams running through
sclerophyllous oak forests; two were recovered from forest soil samples. In
montane forests of the tropical island Hainan, located in the South China Sea
close to Vietnam, six Phytophthora species
were found (Jung et al. 2019).
These
studies are being conducted in the context of scientists discovering numerous
new species of Phytophthora in recent
decades. Since 1999, the number of described species and informally designated
taxa of Phytophthora has tripled. World-renowned
experts Clive Brasier anticipates that between 200 and 600 species of Phytophthora are extant in natural
ecosystems around the world (Jung et al.
2019).
In
the Vietnam survey, P. ramorum was the
most widespread species. While genetic studies indicate ancestral connections to
the four P. ramorum lineages (genetic
strains) introduced to North America or Europe, further studies are under way to clarify these relationships (Jung et al. 2019).
Jung
and colleagues found P. cinnamomi to
be the most common soilborne Phytophthora
species at elevations above 700 m. Two genotypes of the P. cinnamomi A2 mating type are causing epidemics in numerous
natural and managed ecosystems worldwide. There was some evidence that the more
frost sensitive A2 mating type might be spreading into higher altitudes in Vietnam
(Jung et al. 2019).
Most
of the Phytophthora species detected
in the rhizosphere were not associated with obvious disease symptoms. (The
principal exception was the A2 mating type of P. cinnamomi in montane forests in northern Vietnam.) (Jung et al. 2019) This lack of disease greatly reduces the chances of detecting the
oomycetes associated with any plants exported from the region – there are
no symptoms.
Since
southern Yunnan, northern Laos, and the eastern Himalayas belong to the same
biogeographic area those areas might also harbor endemic P. ramorum populations. Further surveys are needed to confirm this
hypothesis (Jung et al. 2019).
Phytophthora
lateralis
– causal agent of Port-Orford cedar root rot – also probably originated in the
area, specifically Taiwan (Vettraino et
al. 2017).
Implications for
phytosanitary measures
Many
of the native Asian forest Phytophthora
species have co-evolved with a variety of tree genera also present in Europe
and North America, including Fagaceae, Lauraceae, Aceraceae, Oleaceae, and
Pinaceae. Numerous examples demonstrate a strong potential that trees in these
families that have not previously been exposed to these Phytophthora species might be highly susceptible. Scientists have
begun an extensive host range study of Phytophthora
species from Asia and South and Central America. One part of this study found
that five Asian Phytophthora species caused
significant rot and loss of fine roots and lateral roots in three European
species of chestnut and oak (Jung et al.
2019).
Other pathogens
Studies by separate groups of scientists have concluded that several beetle-fungus disease complexes are native to this same region.
Sassafras – photo by David Moynihan
Both the laurel wilt fungus Raffaelea lauricola and its primary vector Xyleborus glabratus probably originated in Southeast Asia; there are probably different strains or genetic makeups across their wide ranges. For example, Dreaden et al. 2019 found that the fungus population from Myanmar differed genetically from those found in Japan, Taiwan, and the United States. Others had already expressed concern about the possibility that new strains of R. lauricola might be introduced (Wuest et al. 2017, cited in Cognato et al. 2019).
Cognato
et al. 2019 found that the beetle
occurs in deciduous forests from southern Japan to Northeast India, so genetic
variation across this range is likely. In fact, they have separated the species
X. glabratus into three species. They
found that some of the beetles might thrive at 40o North – the
latitude of central Illinois, Indiana, and Ohio and southern Pennsylvania. The
ability of the vector of laurel wilt disease to spread so far north poses an
alarming threat to sassafras (Sassafras albidum) – which is a major
understory tree in forests of these regions.
It is unknown whether these new species and X. glabratus lineages are associated with different fungal strains. In company with the pathologists cited above, Cognato et al. 2019 warn that preventing introduction of the three beetle species to other regions is prudent. Cognato et al. 2019 point out that if other beetle lineages from the southern extent of their range can tolerate hotter and drier conditions, they might pose a greater risk to host species in the more arid areas of California and Mexico. In addition, Central America is at great risk because of the numerous plant species in the vulnerable Lauraceae found there.
Also from the region are two beetle-fungus combinations killing trees in at least seven botanical families, including maples, oaks, and willows, in southern California. The polyphagous shot hole borer (Euwallacea whitfordiaodendrus) apparently is native to Vietnam (Eskalen et al. 2013) and the closely related Kuroshio shot hole borer (Euwallacea kuroshio) to Japan, Indonesia, and Taiwan (Gomez et al. 2018).
What you can do
Getting APHIS to act
1) communicate concern about the risk to APHIS leadership and ask that the agency take action under its NAPPRA authority
2) communicate the same to intermediaries who can influence APHIS:
State phytosanitary agency – especially through regional plant boards and National Plant Board
Your Congressional representative and senators (especially if one or more serves on Agriculture or Appropriations committee)
Professional societies – American Phytophathological Society, Mycological Society, American Society of Entomologists, Society of American Foresters …
3) communicate the same to university leadership and ask that their lobbyists advocate to USDA
4) communicate the same to the media
2) Research on extent of North American tree species’ vulnerability to the Oomycetes and other associated microorganisms
Jung et al. 2019 say that studies are under
way to identify potential pest-host relationships with important tree species.
However, all the authors are Europeans. Is anyone carrying out tests on North
American trees in the apparently most vulnerable families — Fagaceae,
Lauraceae, Aceraceae, Oleaceae, and Pinaceae?
1)
Communicate with colleagues, scientific societies, APHIS, Agriculture Research
Service, National Institute of Food and Agriculture, and USFS to determine
whether such tests are under way or planned.
2) In those cases where no studies are planned, work with above to initiate them.
Sources
Cognato,
A.I., SM. Smith, Y. Li, T.H. Pham, and J. Hulcr. 2019. Genetic Variability
Among Xyleborus glabratus Populations
Native to Southeast Asia (Coleoptera: Curculionidae: Scolytinae: Xyleborini)
and the Description of Two Related Species. Journal of Economic Entomology XX(XX),
2091, 1 – 11.
Dreaden,
T.J., M.A. Hughes, R.C. Ploetz, A. Black and J.A. Smith. 2019. Genetic Analyses
of the Laurel wilt Pathogen, Raffaelea
lauricola, in Asia Provide Clues on the Source of the Clone that is
Responsible for the Current USA Epidemic. Forests 2019, 10, 37
Eskalen,
A., Stouthamer, R. Lynch, S.C., Twizeyimana, M., Gonzalez, A., and Thibault, T.
2013. Host range of Fusarium dieback and its ambrosia beetle (Coleoptera
Scolytinae) vector in southern California. Plant Disease 97938-951.
Gomez,
D.F., J. Skelton, M.S. Steininger, R. Stouthamer, P. Rugman-Jones, W.
Sittichaya, R.J. Rabaglia, and J. Hulcr1/ 2018. Species Delineation Within the
Euwallacea fornicatus (Coleoptera: Curculionidae) Complex Revealed by
Morphometric and Phylogenetic Analyses. Insect Systematics and Diversity,
(2018) 2(6): 2; 1–11
We
welcome comments that supplement or correct factual information, suggest new
approaches, or promote thoughtful consideration. We post comments that disagree
with us — but not those we judge to be not civil or inflammatory.
Faith Campbell receives award for activism from National Association of State Foresters; 2016
For nearly 30 years I have documented bioinvasion threats and gaps, first in three Fading Forests reports (available here), then in five years of blogging. Here I pull together that information and suggest — in most cases reiterate — steps to address these threats and gaps. I list sources of discussion of the underlying issues – other than my reports and blogs – in references at the end of this blog.
My
first premise is: robust federal leadership is crucial:
The Constitution gives primacy to
federal agencies in managing imports and interstate trade.
Only a consistent approach can
protect trees (and other plants) from non-native pests.
Federal agencies have more
resources than state agencies individually or in any likely collective effort
— despite decades of budget and staffing cuts.
My
second premise is: success depends on a continuing, long-term effort founded on
institutional and financial commitments commensurate with the scale of the threat.
This requires stable funding; guidance by research and expert staff; and engagement
by non-governmental players and stakeholders. Unfortunately, as I discuss
below, funding has not been adequate or stable.
My third premise is that programs’ effectiveness needs to be measured, not just effort (see the NECIS document referenced at the end of the blog).
SPECIFICS
Preventing
new introductions continues to be the most effective action. Mitigating options
decrease and damages increase once a non-native pest has entered the country –
much less become established (see Lovett et
al. 2016 and Roy et al. 2014). I
recognize that preventing new introductions poses an extremely difficult
challenge given the volume and speed of international trade and the strong
economic forces supporting free trade. These challenges have been exacerbated over
several decades by the political zeitgeist – the anti-regulatory ideology, the emphasis
on “collaborating” with “clients” rather than imposing requirements through
regulations. Although the current “America First” policy might reduce import
volumes and therefore reduce the invasive species threat to some extent, the
anti-regulatory stance has only strengthened.
containers at the Port of Long Beach, California
Decades of cutting key agencies’ budgets and personnel are another factor. However, the damage to America’s natural systems is so great that we must try harder to find more effective strategies (See the Fading Forest reports; my previous blogs; Lovett et al. 2016; and APHIS annual reports – e.g., the 2019 report here)
Prevention
Despite adoption and implementation of new international and national regulations to stem pest introductions, introductions continue – although probably at a lower level than would otherwise be the case. Delays in adoption of regulations (documented in Fading Forests II and III and my two recent 30-years-in-review blogs have facilitated damaging introductions and spread.
Solutions
Stakeholders press USDA
leadership to initiate rules intended to strengthen phytosanitary protection and
expedite their completion
APHIS promote and facilitate
analysis of current programs and policies by non-agency experts to ensure the
agency is applying most effective strategies (Lovett et al. 2016).
Adoption
of insufficiently protective regulations (documented in FFII, FFIII, two
30-years-in-review blogs) – adopted in part because APHIS is trying to
“balance” trade facilitation and phytosanitary protection – has further
contributed to damaging pests’ introduction and spread.
Solutions:
Boost
priority of preventing pest introductions by amending the Congressional finding
in the Plant Protection Act [7 USC 7701(3)] as follows
Existing language: “[I]t is the
responsibility of the Secretary [of Agriculture] to facilitate exports, imports
and interstate commerce in . . . commodities that pose a risk of harboring
plant pests or noxious weeds in ways that
will reduce, to the extent practicable, as determined by the Secretary, the
risk of dissemination of plant pests and noxious weeds .… “
Amend to read as follows: “…. in ways that will reduce prevent, to the greatest extent practicable feasible, as determined by the Secretary, …” [emphasis added]
Adopt several actions to
strengthen phytosanitary protections at the point of origin (Lovett et al. 2016)
Expand
pre-clearance partnerships — as authorized for plants under Q-37 regulations
and ISPM-36
Expand
sentinel tree programs
Promote
voluntary substitution of packaging made from materials other than solid wood.
APHIS
doesn’t use the enforcement powers that it has under Plant Protection Act (see
several of my past blogs)
Solutions:
CBP inspectors search for pests in a pallet; CBP photo
APHIS follow the lead of Customs and Border Protection and begin penalizing importers on the first instance of their wood packaging not being in compliance with ISPM#15 (see blog here).
APHIS prohibit use of wood packaging by countries and importers of categories of imports that – over the 13 years since implementation – have developed a record of frequent violations of ISPM#15.
APHIS use its authority per revised Q-37 regulations to negotiate with countries that export plants to the U.S. to establish “integrated measures” programs aimed at minimizing the risk of associated pests being transported to the U.S.
APHIS use its authority per revised Q-37 to place in the “Not Authorized for Import Pending Pest Risk Assessment (NAPPRA) “limbo” category genera containing North American “woody” plants (see Roy et al. 2014; Lovett et al. 2016).
Spread within the
U.S.
The
United States lacks a coordinated system to prevent pest spread within the
country (see Fading Forests III Chapter 5). Even our strictest methods, like APHIS’s
quarantines regulating interstate movement of goods, have failed to curtail
spread of significant pests. The most obvious example is the emerald ash borer.
The regulations governing movement of the sudden oak death pathogen in the nursery trade have also failed: there have been periodic outbreaks in which the pathogen has been spread to nurseries across the country. Between 2003 and 2011, a total of 464 nurseries located in 27 states tested positive for the pathogen, the majority as a result of shipments traced from infested wholesalers. In 2019, plants exposed to the pathogen were again shipped to 18 states; eight of those states have confirmed that their plant retailers received infected plants (see my blog from summer here).
Another
serious gap is the frequent failure of APHIS and states to adopt official
programs targetting bioinvaders that will be difficult to control because of
biological characteristics or cryptic natures – even when severe impacts are
demonstrated. Recent examples include the laurel wilt disease complex, goldspotted
oak borer, polyphagous and Kuroshio shot hole borers and associated pathogens,
and even the spotted lanternfly (although the last has received significant
funds from APHIS.)
redbay killed by laurel wilt disease, Georgia; photo by Scott Cameron
Solutions:
APHIS apply much more stringent
regulations to interstate movement, based on a heightened priority for
prevention in contrast to facilitating interstate trade. E.g., prohibit nurseries on the West Coast from shipping P. ramorum hosts to states where the pathogen
is not established.
APHIS encourage states to adopt
quarantines and regulations aimed at preventing spread of invasive pests to
regions of the state that are not yet infested. For example, the sudden oak
death pathogen in California and Oregon; the borers in southern California.
APHIS abandon plans to deregulate
emerald ash borer and step up its support for state regulations on firewood.
APHIS stop dumping pests it no
longer wants to regulate onto the states through the “Federally Recognized
State Manage Phytosanitary (FRSMP) program”.
APHIS revise its policies so that
the “special needs exemption” [7 U.S.C. 7756] actually allows states to adopt
more stringent regulations to prevent introduction of APHIS-designated
quarantine pests (see Fading Forests III Chapter 3).
To help fill the gaps, the states are trying to coordinate their regulations in some important areas. The most advanced example is the voluntary Systems Approach to Nursery Certification, or SANC program. APHIS has supported this initiative, including by funding from the Plant Pest and Disease Management and Disaster Program (see below). However, it is a slow process; USDA funds first became available in 2010. The states are trying to coordinate on firewood, but we don’t yet know what the process will be.
Funding shortfalls (See the three Fading Forests
reports, my blogs about appropriations)
Increase APHIS’ access to emergency
funds from the Commodity Credit Corporation by
amending the Plant Protection Act [7 U.S.C. 7772 (a)] to include this
new definition of “emergency”:
the term “emergency” means any
outbreak of a plant pest or noxious weed which directly or indirectly threatens
any segment of the agricultural production of the United States and for which
the then available appropriated funds are determined by the Secretary to be
insufficient to timely achieve the arrest, control, eradication, or prevention
of the spread of such plant pest or noxious weed.
Although APHIS has the most
robust prevention program of any federal agency, its funding is still
inadequate. Stakeholders should lobby the Congress in support of higher annual
appropriations.
The Plant
Pest and Disease Management and Disaster Program (now under Section 7721 of the
Plant Protection Act) has provided at least $77 million for tree-pest
programs (excluding NORS-DUC & sentinel plant programs and other programs)
since FY 2008. Much useful work has been carried out with these funds. However,
these short-term grants cannot substitute for stable, long-term funding. I
reiterate my call for stakeholders to lobby the Congress to provide larger
appropriations to the APHIS Plant Protection program and Forest Service Forest
Health Protection and Research programs.
Long-term Responses
to Bioinvasive Challenge
More stakeholders are advocating raising the priority of – and providing adequate resources to – such long-term solutions as biocontrol and breeding trees resistant to pests and restoring them to our forests. Advocates include the state forestry agencies of the Northeast and Midwest, some non-governmental organizations, some academics, and individual USFS scientists. One effort resulted in inclusion of language in the 2018 Farm Bill (see blog here) – although this approach has apparently run into a dead end. The new emphasis on breeding has so far not been supported by agency or Congressional leaderships.
test planting of an American chestnut bred to be resistant to chestnut blight
Solutions:
USFS convene workshop of the
federal, state, National Academy, academic, and NGO groups promoting resistance
breeding in order to develop consensus on priorities and general structure of program.
Explicitly include evaluation of the
CAPTURE Project’s (see blog here) efforts to
set priorities to guide funding allocations and policies; and proposals for
providing needed supportive infrastructure – facilities, trained staff in
various disciplines. (See my blogs here.)
Report results of meeting to USDA
leadership, Congress, and stakeholders
Then ensure implementation of the
accepted approach by both Research and Development and Forest Health Protection
programs. Include provisions to provide sustainable funding.
These proposed actions still do not address ways to correct the provisions of the international phytosanitary agreements (World Trade Organization and International Plant Protection Convention) that complicate – or preclude – efforts to prevent introduction of pests currently unknown to science. This issue is discussed in Fading Forests II. A current example is beech leaf disease (described here).
Continuing
inadequate engagement by stakeholders
Most
constituencies that Americans expect to protect our forests don’t press
decision-makers to fix the problems I have identified above: inadequate
resources, weak and tardy phytosanitary measures. Some of these stakeholders
are other federal agencies, or state agencies – or their staffs. They face
restrictions on how “political” they can be. But where are the professional and scientific associations,
representatives of the wood products industry, forest landowners, environmental NGOs and their funders, urban
tree advocates Efforts by me, USDA, and others to better engage these groups
have had disappointing results.
As
I have documented, groups of USFS scientists have made several attempts to
document the extent of invasive species threats and impacts and to set
priorities. So far, they have not gained much traction. Another USFS attempt,
Poland et al. in press, will appear
at the end of the year. Will this be more successful?
I
detect growing attention to educating citizen scientists for early detection;
but if there is an inadequate – or no – official response to their efforts
won’t people become discouraged?
SOURCES
Lovett,
G.M., M. Weiss, A.M. Liebhold, T.P. Holmes,
B. Leung, K.F. Lambert, D.A. Orwig, F.T. Campbell, J. Rosenthal, D.G. McCullough,
R. Wildova, M.P. Ayres, C.D. Canham, D.R. Foster, SL. Ladeau, and T. Weldy.
2016. NIS forest insects and pathogens in the US: Impacts and policy options. Ecological
Applications, 26(5), 2016, pp. 1437–1455
National Environmental Coalition on Invasive Species “Tackling the Challenge.”
Poland,
T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019),
Invasive Species in Forests and Grasslands of the United States: A
Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. (in press).
Roy,
B.A., H.M Alexander, J. Davidson, F.T Campbell, J.J Burdon, R. Sniezko, and C.
Brasier. 2014. Increasing forest loss worldwide from P&Ps requires new
trade regulations. Front Ecol Environ 2014; 12(8): 457–465
It
is widely recognized that invasions of non-native species occur as a
consequence of international trade (see Seebens et. al. 2017 – full citations at the end of this blog). This is as
true for non-native forest pests as for any other bioinvader – see Aukema et al. 2010; Liebhold et al. 2012, Lovett et al. 2016. In fact, gross domestic product – as an indicator of levels
of trade — is a better predictor of the number of forest pest invasions in a
given country than the country’s amount of forested land (Roy et al. 2014).
shipping containers at port of Long Beach, California
As I noted in my previous blog, I began studying and writing about the threat to North America’s forests from non-native insects and pathogens in the early 1990s. I reported my analyses of the evolving threat in the three “Fading Forests” reports – coauthored by Scott Schlarbaum – in 1994, 2003, and 2014. These reports are available here.
I document here that both introduction
and spread of pests within the country have continued apace. While significant
efforts have been made to prevent introductions (described briefly under the
“Invasives 101” tab of the CISP website), they have fallen short. As I noted in
Fading Forests III, programs aimed at preventing spread of pests within the
country remain fragmented and often are unsuccessful.
The Challenge: Huge Volumes of goods are moving, providing
opportunities for pests
Since 1990, volumes of imported goods more than quintupled. Within the U.S., a total of 17,978 million tons of goods were transported in 2015; 10,776 million tons of this total by truck. About one-third of this total – 5,800 million tons – was moved farther than 250 miles. These vehicles moved on a public roads network of 4,154,727 miles (US DOT FFA). Consequently, once a pest enters the U.S., it can be moved quickly into every corner of the country.
Introductions
By and large,
establishment of tree-killing pests has occurred at a fairly steady rate of
about 2.5 per year, with “high-impact” insects and pathogens accumulating at
0.43 per year (Aukema et al. 2010). Since
introductions did not rise commensurately with rising import volumes, Lovett et al. (2016) concluded that the
recently adopted policies for preventing introductions referenced above are
having positive effects but are insufficient to reduce the influx of pests in
the face of ever-growing global trade volumes. The study’s authors went on to
say that absent more effective policies, they expect the continued increase in
trade will bring many new establishments of non-native forest pests.
One group of forest
pests did not enter at a steady rate, but rather entered at a higher rate since
1985 – wood-boring insects. Experts concluded that the increase probably
reflected increases in containerized shipping (Lovett et al. 2016). At the global level, the rate of fungal invasions has
also recently been reported to be increasing rapidly (Roy et al. 2014).
Asian longhorned beetle
Geography of trade
patterns also matters. Opening of trade with China (in 1979) offered
opportunities for pests from a new source country which has a similar climate
and biology. Roy et al. describe the importance of phylogenetic
relatedness of pests and of tree hosts in explaining tree species’ vulnerability
to introduced pests. The most vulnerable forests are those made up of species
similar to those growing in the source of the traded goods – i.e., the temperate forests of the
northeastern U.S. – when goods are imported from similar forested areas of
Europe and Asia. Chinese-origin wood-boring pests began to be detected around
1990. This already short interval probably underestimates how quickly pests
began arriving; detection methods were poor in those years, so a pest was often
present for close to a decade before detection.
Between 1980 and 2016, at least 30 non-native species of wood- or bark-boring insects in the Scolytinae / Scolytidae were newly detected in the United States (Haack and Rabaglia 2013; Rabaglia et al. 2019). Over the same period, approximately 20 additional tree pests were introduced to the continental states (Wu et al. 2017; Digirolomo et al. 2019; R. Haack, pers. comm.) plus about seven to America’s Pacific islands. Not all of the new species are highly damaging, but enough are. See my previous blog here.
Many of the tree-killing
pests were probably associated with pathways other than wood packaging. These
include 6 of the 7 Agrilus species, sudden oak death pathogen, three pests of palm trees, the
spotted lanternfly, beech leaf disease; and the pests introduced to America’s
Pacific Islands.
HIGH-RISK
PATHWAYS OF INTRODUCTION
Already
in the 1990’s it was evident that better preventing pest introductions would
depend on shutting down the variety of pathways by which they move around the
world. At that time, attention focused on imports of
logs and nursery stock (nursery stock makes up one component of a broader
category called by phytosanitary agencies “plants for planting”). Both logs and
“plants for planting” had well-established histories of transporting pests and
import volumes were expected to grow. We have since learned that there are many
more pathways!
Plants for Planting
Imports of “plants for planting” (phytosanitary agencies’ term, which encompasses nursery stock, roots, bulbs, seeds, and other plant parts that can be planted) have long been recognized as a dangerous pathway for introduction of forest pests. For example, this risk was the rationale for adopting the 1912 Plant Quarantine Act. Charles Marlatt, Chairman of USDA’s Federal Horticultural Board (see “Then and Now” in Fading Forests III here), wrote about the risk in National Geographic in April 1911 (urging adoption of the 1912 law) and again in August 1921. See also Brasier (2008), Roy et al. (2014), Liebhold et al. (2012), Jung et al. (2016).
Japanese cherry trees being burned because of scale infestation January 28, 1910; Agriculture Research Service
Of the 91 most
damaging non-native forest pest species in the U.S. (Guo et al. 2019), about 62% are thought to have entered North America
with imports of live plants. These include nearly all the sap-feeding insects,
almost 90% of the foliage-feeding insects, and approximately half of the
pathogens introduced during the period 1860-2006 (Liebhold et al. 2012). Specific examples include chestnut blight, white pine
blister rust, Port-Orford-cedar root disease, balsam woolly adelgid, hemlock
woolly adelgid, beech scale, butternut canker, dogwood anthracnose, and sudden
oak death. In more recent years, introductions via this pathway possibly
include ‘ōhi‘a
rust, rapid ‘ōhi‘a death pathogens, and beech leaf disease. The gypsy
moth, while a foliage feeder, was not introduced via imports of live plants.
The
APHIS annual report for 2018 reported that in that year we imported 18,502
shipments containing more than 1.7 billion
plant units (plants, bulbs, in vitro materials, etc.).
Liebhold et al. 2012, relying on 2009 data, found that about 12 percent of incoming plant shipments had symptoms of pests – a rate more than 100 times greater than that for wood packaging. Worse, a high percentage of the pests associated with a shipment of plants is not detected by the federal inspectors. The meaning of this finding is unclear because the study did not include any plant genera native to temperate North America and APHIS points out that infestation rates varied considerably among genera in the study. However, APHIS has not conducted its own analysis to document the “slippage rate” on imports of greatest concern to forest conservationists, i.e., imports of woody plants. I provide details on pests detected on imports of woody plants in recent in my blog here.
Clearly the risk of pest introductions continued at least until recently. I reviewed an APHIS database listing pests newly detected in the country during the period 2009-2013. I concluded that approximately 37 of the 90 “new” pests listed in the database (viruses, fungi, aphids and scales, whiteflies, mites) were probably introduced via imports of plants, cuttings, or cut foliage or flowers. I discussed these matters in greater detail here.
Adoption of a new regulatory regime governing imported plants for planting (Q-37 regulation) in 2018 is too recent to for us to see its impact. But the new regulation sets up a process under which APHIS can impose more protective regulations on specific types of plants or plants from certain countries of origin to counter a perceived concerning level of risk. Until APHIS begins activating its new powers by negotiating more protective regulations governing plant imports from high-risk sources, it seems unlikely there will be any meaningful change in the introduction rates.
Crates, Pallets,
and Other Forms of wood packaging (solid wood packaging, or SWPM)
Recognition
of the risk associated with wood packaging is much more recent. In 1982, a USDA
risk assessment concluded that the wood boring insects found in crates and
pallets were not of great concern (USDA APHIS and Forest Service, 2000).
However, contradictory indications were quickly documented – including from
APHIS’ own port interception data – which the agency began collecting in 1985. Over
the 16-year period 1985-2000, 72%
of
the 6,825 bark beetles (Scolytidae)
intercepted by APHIS were found on SWPM (Haack 2002). Cerambycids (longhorned
beetles) and buprestids (jewel beetles) make up nearly 30% of insects detected
in wood packaging over the last 30 years (Haack et al. 2014).
Detection
of outbreaks of the Asian longhorned beetle and other woodborers in the
mid-1990s made it clear that wood packaging was, indeed, a high-risk pathway.
Of
the 91 most damaging non-native pest species in the US, 30% probably arrived
with wood packaging material or other wood products (Liebhold et al. 2012). This group includes many
of the most damaging pests, the deadly woodborers – Asian longhorned beetle,
emerald ash borer, redbay ambrosia beetle,
possibly the polyphagous and Kuroshio shot hole borers.
CBP agents inspecting a pallet
As noted above, introductions of wood borers have risen in recent decades, widely accepted as associated with the rapid increase in containerized shipping after 1980. In 2009 it was estimated that 75% of maritime shipments were packaged in crates or pallets made of wood (Meissner et al. 2009). A good history of the global adoption of containerized shipping is Levinson, M. The Box: How the Shipping Container Made the World Smaller and the World Economy Bigger (Princeton University Press 2008)
The simultaneous
opening of trade with China (in 1979) offered opportunities for pests from a
new source country which has a similar climate and biology. Chinese-origin wood-boring
pests began to be detected around 1990. This already short interval probably underestimates
how quickly pests began arriving; detection methods were poor in those years,
so a pest was often present for close to a decade before detection.
I have already documented numerous times that, despite the U.S.’ implementation of the International Standard of Phytosanitary Measures (ISPM) #15 in 2006, live quarantine pest woodborers continue to enter the U.S. in wood packaging. The best estimate is that 0.1% of wood packaging entering the United States is infested with wood-borers considered to be quarantine pests (Haack et al. 2014). More than 22 million shipping containers entered the U.S. via maritime trade in 2017 (US DoT). As noted, an estimated 75% of sea-borne containers include wood packaging. Applying the 0.1% estimate to these figures results in an estimate that as many as 17,650 containers per year (or 48 per day) transporting tree-killing insects enter the U.S.
Over a period of
nine years – Fiscal Years 2010 through 2018 – U.S. Customs and Border
Protection (CBP) detected more than 28,600 shipments with wood packaging that
did not comply with ISPM#15 (Harriger presentations to the annual meetings of
the Continental Dialogue on Non-Native Forest Insects and Diseases). While most
of the non-compliant shipments were wood packaging that lacked the required
mark showing treatment per ISPM#15,
in 9,500 cases the wood packaging actually harbored a pest in a regulated
taxonomic group.
Disturbingly, 97%
of the shipments that U.S. CBP found with infested wood packaging bear the
ISPM#15 mark certifying that wood had been fumigated or heat-treated (Harriger
2017). CBP inspectors tend to blame this on widespread fraud in use of the mark.
On the other hand, one study found that larvae can survive both treatments –
although the frequency of survival was not determined. It was documented that twice
as many larvae reared from wood treated by methyl bromide fumigation survived
to adulthood than larvae reared from heat-treated wood; the reason is unclear
(Nadel et al. 2016).
The APHIS’ record
of interceptions for the period FYs 2011 – 2016 contained 2,547 records for
insect detections on wood packaging. The insects belonged to more than 20
families. Families with the highest numbers of detections were Cerambycids –
25% of total; Curculionidae – 23% (includes Dendroctonus,
Ips, Orthotomicus, Scolytinae, Xyleborus, Euwallacea); Scolytidae – 17% (includes
true weevils such as elm bark beetles); Buprestids – 11%; and Bostrichidae –
3%. Not all of the insects in these groups pose a threat to North American
plant species.
One encouraging data point is that since 2010, there have been no detections of species of bark and ambrosia beetles new to North America in the traps deployed by the USDA Forest Service Early Detection and Rapid Response program (Rabaglia 2019). The 2014 recognition of the Kuroshio shothole borer apparently did not result from this trapping program.
There have been several changes in the wood packaging standard and its implementation by CBP since 2009, the year Haack et al. 2014 analyzed the “pest approach rate”. APHIS has not carried out a study to determine whether these recent changes have reduced the approach rate below Haack’s estimate of 0.01%. Consequently, we do not know whether these changes have reduced the risk of pest introductions.
Other Pathways That
Transport Fewer Pests – Some of Which Have High Impacts
Insects
that attach egg masses to hard surfaces can be transported by ship
superstructures, containers, and hardsided cargoes such as cars, steel beams,
and stone. While relatively few species have been moved in this way, some have
serious impacts. The principal examples are the gypsy moths from Asia, which
feed on 500 species of plants (Gibbon 1992).
The United States and Canada have a joint program – under the auspices of the North American Plant Protection Organization (see RSPM #33) aimed at preventing introduction of species of Asian gypsy moths. The NAPPO standard originally went into force in March 2012. Under its terms, ships leaving ports in those countries during gypsy moth flight season must be inspected and cleaned before starting their voyage.
Gypsy moth populations rise and fall periodically; it is much more likely that egg masses will be attached to ships during years of high moth population densities. These variations are seen in U.S. and Canadian detection reports – as reported here.
While most AGM detections are at West Coast ports, [here; and here] the risk is not limited to that region. AGM have been detected at Wilmington, NC; Baltimore, MD; Charleston, SC; Savanna and Brunswick, GA; Jacksonville, FL; New Orleans, LA; Houston and Corpus Christi, TX; and even McAlester, OK.
Nor is the risk limited to the ships themselves. In 2014, more than 500 Asian gypsy moth egg masses were found on four shipments of imported steel slabs arriving at ports on the Columbia River in Washington.
Between 1991 and 2014, AGM was detected and eradicated
on at least 20 occasions in locations across the United States (USDA AGM pest
alert). Additional outbreaks have been discovered and eradication efforts
undertaken in more recent years.
A second example is
the spotted lanternfly (SLF) (Lycorma delicatula), which was first
detected in southeast Pennsylvania in autumn 2014. It is native to Asia; it is
believed to have entered the country as egg masses on imported stone.
While SLF is
clearly a pest of agriculture – especially grapes and tree fruits – its
importance as a forest pest is still unclear. Many native forest trees appear
to be hosts during the insect’s early stages, including maples, birches,
hickories, dogwoods, beech, ash, walnuts, tulip tree, tupelo, sycamore, poplar,
oaks, willows, sassafras, basswood, and elms. Adult lanternflies strongly
prefer the widespread invasive species tree of heaven (Ailanthus altissima).
As of August 2019, SLF was established in parts of five states: Delaware, Maryland, New Jersey, Pennsylvania, and Virginia. It was detected as having spread to a 14th county in Pennsylvania; five new counties in New Jersey. APHIS is working with state departments of Agriculture in these states, as well as supporting surveys in New York, North Carolina, and West Virginia (USDA APHIS DA-2019-20, August 7, 2019). Apparently the detections of a few adults – alive or dead – in Connecticut and New York had not evolved into an outbreak. See description and map here.
Imports of logs – roundwood – seem inherently risky. Certainly Dutch elm disease was introduced via this pathway. However, there have been few pest introductions linked to this pathway in recent years, probably because we import most of our unprocessed lumber from Canada. (I provide considerable data on U.S. roundwood imports in Fading Forests III here.)
Decorative items and furniture made of unprocessed wood certainly have the potential to transport significant pests (USDA APHIS 2007). Examples include boxes and baskets; wood carvings; birdhouses; artificial Christmas trees or other plants; trellises; lawn furniture. To date, apparently, no high-impact pest has been introduced via this pathway, although pests intercepted on shipments have included Cerambycids from Asia, e.g., velvet longhorned beetle and here.
Alarmed by high numbers of infested shipments from China, APHIS first suspended imports of such items temporarily; then adopted a regulation (finalized in March 2012 – USDA APHIS 2012).
APHIS
has not taken action to prevent introductions on such items imported from other
countries – although the North American Plant Protection Action adopted a
regional standard making the case for such action and outlining a risk-based
approach (NAPPO RSPM#38).
Snails on Shipping
Containers
Snails have been
detected on shipping containers and wood packaging for decades. In 2015, APHIS
stepped up its efforts to address this risk through bilateral negotiations with
Italy and launching regional and international efforts to develop guidance for
ensuring pest-free status of shipping containers (Wendy Beltz, APHIS, presentation
to National Plant Board, 2018 annual meeting).
SPREAD
WITHIN THE UNITED STATES
Major pathways for human-assisted spread of pests within the country are sales of plants for planting, movement of unprocessed wood – especially firewood, and hitchhiking on transport vehicles. Since most forest pests are not subject to federal quarantine, any regulatory programs aimed at preventing spread depend on cooperation among the 50 states. None of these pathways is regulated adequately to prevent pests’ spread. See Chapter 5 of Fading Forests III here.
And since neither
federal nor state agencies do significant enforcement of existing regulations,
preventing spread often depends upon pest awareness of, and voluntary
compliance by, individuals and companies.
Even pests subject to a federal quarantine are not prevented from spreading. Plants exposed to the sudden oak death pathogen were shipped to 18 states in spring 2019.
SOD-infected rhododendron plant; Indiana Department of Natural Resources
A collaborative effort by the nursery industry, APHIS, and states (Systems Approach to Nursery Certification, or SANC) is striving to close gaps linked to the standard practice of inspecting plants at the time of shipping, but full implementation of this voluntary program is still years away.
Transport of firewood has been responsible for movement of pests both short distances, e.g., goldspotted oak borer in southern California; and long distances – e.g., emerald ash borer to Colorado. APHIS attempted to develop a certification program but the industry was unable to put one together (see Chapter 5 of Fading Forests III). Current federal and state regulations of firewood are tied to the emerald ash borer quarantine, which APHIS has proposed to terminate. Wood for turning and woodworking has also been linked to movement of pests, e.g., walnut twig beetle/thousand cankers disease from the west to Pennsylvania.
emerald ash borer
Truck transport of a
variety of goods has transported European gypsy moths from the infested areas
in the east to the west coast. Transport of stone probably moved spotted
lanternfly from southeastern Pennsylvania to Winchester, Virginia.
SOURCES
Aukema, J.E., D.G. McCullough, B. Von
Holle, A.M. Liebhold, K. Britton, & S.J. Frankel. 2010. Historical
Accumulation of Nonindigenous Forest Pests in the Continental United States.
Bioscience. December 2010 / Vol. 60
No. 11
Brasier, C.M. 2008. The biosecurity
threat to the UK and global environment from international trade in plants. Plant Pathology (2008) 57, 792-808
Bray, A.M., L.S. Bauer, T.M. Poland,
R.A. Haack, A.I. Cognato, J.J. Smith. 2011. Genetic analysis of emerald ash
borer (Agrilus planipennis Fairmaire)
populations in Asia and North America. Biol. Invasions (2011) 13:2869-2887
Gibbon,
A. 1992. “Asian Gypsy Moth Jumps Ship to United States.” Science.
Vol. 235. January 31, 1992.
Haack R. A. and J.F. Cavey. 1997.
Insects Intercepted on Wood Articles at United States Ports-of-Entry and Two
Recent Introductions: Anoplophora
glabripennis and Tomicus piniperda.
In press in International forest insect workshop proceedings, 18 – 21
August 1997, Pucon, Chile. Corporacion National Forestal, Santiago, Chile.
Haack, R.A., F. Herard, J. Sun, J.J.
Turgeon. 2010. Managing Invasive Populations of Asian Longhorned Beetle and
Citrus Longhorned Beetle:A Worldwide Perspective. Annu. Rev. Entomol. 2010.
55:521-46.
Haack,
R.A. and R.J. Rabaglia. 2013. Exotic Bark and Ambrosia Beetles in the USA:
Potential and Current Invaders. CAB International 2013. Potential Invasive
Pests of Agricultural Crops (ed. J. Peña)
Haack R.A., Britton K.O., Brockerhoff, E.G., Cavey, J.F.,
Garrett., L.J., 2014. Effectiveness of the International Phytosanitary Standard
ISPM No. 15 on Reducing Wood Borer Infestation Rates in Wood Packaging Material
Entering the United States. PLoS ONE 9(5): e96611.
doi:10.1371/journal.pone.0096611
Haack,
R.A., F. H´erard, J. Sun, and J.J. Turgeon. 2010. Managing Invasive Populations
of Asian Longhorned Beetle and Citrus Longhorned Beetle: A Worldwide
Perspective. Annu. Rev. Entomol. 2010. 55:521–46
Harriger, K. Department of Homeland Security Bureau of
Customs and Border Protection, presentation to the Continental Dialogue on
Non-Native Forest Insects and Diseases, November 2017.
Jung T, Orlikowski L, Henricot B, et al. 2016. Widespread
Phytophthora infestations in European nurseries put forest, semi-natural and
horticultural ecosystems at high risk of Phytophthora diseases. Forest
Pathology 46: 134–163.
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L.Parke, and K.O Britton.
2012. Live plant inports: the major pathway for forest insect and pathogen invasions of the US. Frontiers in
Ecology.
Lovett, G.M., M. Weiss, A.M. Liebhold, T.P. Holmes, B.
Leung, K.F. Lambert, D.A. Orwig, F.T.
Campbell, J. Rosenthal, D.G. McCullough, R. Wildova, M.P. Ayers, C.D. Canham,
D.R. Foster, S.L. LaDeau, and T. Weldy. 2016.
Non-native forest insects and pathogens in the United States: Impacts
and policy options. Eological
Applications, 26(5) pp. 1437-1455.
Meissner, H., A. Lemay, C. Bertone, K. Schwartzburg, L.
Ferguson, and L. Newton. 2009. EVALUATION OF PATHWAYS FOR EXOTIC PLANT PEST
MOVEMENT INTO AND WITHIN THE GREATER CARIBBEAN REGION. Caribbean Invasive
Species Working Group (CISWG) and Plant Epidemiology and Risk Analysis
Laboratory (PERAL) / CPHST. June 4, 2009
Morin, R. presentation at Northeastern Forest Pest Council 81st Annual
Meeting, March 12 – 14, 2019, West
Chester, Pennsylvania
Nadel, N., S. Myers, J. Molongoski, Y.
Wu, S. Linafelter, A. Ray S. Krishnankutty, and A. Taylor. 2016.
Identificantion of Port Interceptions in Wood Packaging Material: Cumulative
Progress Report, April 2012 – August 2016
Rabaglia,
R.J., A.I. Cognato, E. R. Hoebeke, C.W. Johnson, J.R. LaBonte, M.E. Carter, and
J.J. Vlach. 2019. Early Detection and Rapid Response. A Ten-Year Summary of the
USDA Forest Service Program of Surveillance for Non-Native Bark and Ambrosia
Beetles. American Entomologist Volume 65, Number 1
Roy, B.A., H.M. Alexander, J. Davidson,
F.T. Campbell, J.J. Burdon, R. Sniezko, and C. Brasier. 2014. Frontiers in Ecology 12(8): 457-465
U.N. Food and Agriculture Organization International
Plant Protection Convention. 2012. International Standards for Phytosanitary Meaures
No. 36 Integrated Measures for Plants for planting. Rome. Online at
https://www.ippc.int/ Accessed December
7, 2012.
United States Department of Agriculture
Animal and Plant Health Inspection Service and Forest Service, 2000. Pest Risk
Assessment for Importation of Solid Wood Packing Materials into the United
States.
United States Department of Agriculture Animal and
Plant Health Inspection Service 2009. Proposed Rule Importation of plants for
planting: establishing a category of plants for planting not authorized for
importation pending pest risk assessment. Federal
Register 74(140): 36403-36414 July 23, 2009.
United
States Department of Agriculture Animal and Plant Health Inspection Service 7
CFR Parts 318, 319, 330, 340, 360, and 361. Federal Register Rules and
Regulations Vol. 83, No. 53. Monday, March 19, 2018
United States Department of Agriculture Animal and
Plant Health Inspection Service 2007. Pests and
mitigations for manufactured wood décor and craft products from China for
importation into the United States. Revision 6. July.
United States Department of Agriculture Animal and
Plant Health Inspection Service. 2012. Importation of wooden handicrafts from
China. Final rule. Federal Register 77(41):
12437-12444. March 1. Online at http://www.gpo.gov/fdsys/pkg/FR-2012-03-01/pdf/2012-4962.pdf.
Accessed August 2, 2013.
United
States Department of Transportation Bureau of Transportation Statistics Freight
Facts and Figures
United States Department of Transportation, Maritime
Administration, U.S. Waterborne Foreign Container Trade by U.S. Customs Ports
(2000 – 2017) Imports in Twenty-Foot Equivalent Units (TEUs) – Loaded
Containers Only
Williams, L.H. and J.P. La Fage. 1979.
Quarantine of Insects Infesting Wood in International Commerce. in J.A.
Rudinksy, ed. Forest Insect Survey and Control Fourth Edition 1979
Wu,Y.,
N.F. Trepanowski, J.J. Molongoski, P.F. Reagel, S.W. Lingafelter, H. Nadel1,
S.W. Myers & A.M. Ray. 2017. Identification of wood-boring beetles
(Cerambycidae and Buprestidae) intercepted in trade-associated solid wood
packaging material using DNA barcoding and morphology Scientific Reports 7:40316