Photo of infested cactus at Cabo Rojo National Wildlife Refuge, Puerto Rico. Taken August 20, 2018 by Yorelyz Rodríguez-Reyes
For 15 years I have maintained a profile of the Harrisia cactus mealybug Hypogeococcus pungensbecause it threatens columnar cacti in the subfamily Cactoideae from the Caribbean basin and possibly in the American Southwest and Mexico. My recent attempts to clarify the current situation have been frustrated by the apparent collapse of funding support for scientists trying to conserve these cactus species.
The dry regions of the Caribbean Islands are home to about 100 native cacti, 75% of which are endemic. According to the Center for Plant Conservation, 20 species are listed as threatened by the IUCN. Puerto Rico specifically is home to 14 cactus species, at least three of which are endemic.
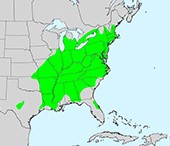
How and when the mealybug first invaded the Caribbean and North America is unknown. The presence of the insect now known as H. pungens Hyp-C on Puerto Rico was detected in the Guánica Commonwealth Forest and Biosphere Reserve on the island’s southern coast in 2005 (Zimmerman et al. 2010). However, the actual introduction probably occurred about ten years earlier, in about 1996 (Poveda-Martinez et al. 2022). See map of locations below.
In the 20 years since then, the mealybug has spread across the island’s dry districts. By 2010, it was estimated to be present on about 1,400 km2. By 2014 – nine years after detection — the mealybug had reached the small island of Caja de Muertos. The most recent survey of which I am aware (date unclear) detected the mealybug on 268 out of 445 cactus plants examined (60%) in 12 out of 39 sites examined (Poveda-Martinez et al. 2022). The mealybug is also killing native cacti on the nearby U.S. Virgin Islands (Poland et al. 2019), although I have found no data on this invasion or its impact.
Below – columnar cacti on St. John, US Virgin Islands; photos by F.T. Campbell
H. pungens Hyp‑C threatens seven of 14 native cactus species in Puerto Rico. Three of the cacti are endemic; two are federally listed as endangered species: Harrisia portoricensis and Leptocereus grantianus (USDA ARS). Since the mealybug’s detection in Puerto Rico, it has caused extensive damage to Pilosocereus royenii (Royen’s tree cactus), Leptocereus qaudricostatus (pitaya), Melocactus intortus (turk’s cap), and an introduced cultivar, Cereus hexagonus. It has caused minor damage to Stenocereus fimbriatus (Zimmerman et al. 2010). These cacti provide food or shelter for endemic bats, birds, moths and other pollinators (Segarra and Ramirez; USDA ARS).
The insect’s attack promotes abnormal gall-like growth on the stem and deformed flowers. These deformations severely affect infested plants’ reproduction and eventually survival (Poveda-Martinez et al. 2022).
Biological Control
When the mealybug was first detected Commonwealth and federal agencies tried to counter it. A search for possible biocontrol agents in the insect’s native range in Argentina began the 2010. While no funds have ever been appropriated for this activity, for several years the U.S. Department of Agriculture supported the work by allocating funds to the Agriculture Research Service Insect Behavior and Biocontrol laboratory in Gainesville, Florida, from broader programs. Dr. Stephen Hight took the lead, working with colleagues in South America. According to Dr. Hilda Diaz-Soltero, then a USDA official, these funds came primarily from the USDA Invasive Species Coordination Program and APHIS Eastern Region. In fiscal years 2017 and 2019, an additional ~$550,000 came in the form of grants under APHIS’ Plant Pest and Disease Management and Disaster Prevention Program. Link Scientists at the Center for Excellence in Quarantine and Invasive Species at the University of Puerto Rico devoted at least a decade to the search.
Scientists focused on two parasitoid wasps, Anagyrus cachamai and A. lapachosus (Hymenoptera: Encyrtidae). A third candidate, the predator Hyperaspis conclusa, was also assessed (Aguire et al.).
Research on the mealybug-wasps interaction uncovered troubling patterns. First, it has long been known that some mealybugs believed to be belong to the species Hypogeococcus pungens feed on columnar cacti while others feed on plants in two unrelated families, Amaranthaceae and Portulacaceae (USDA ARS; Zimmerman et al. 2010). Would the introduced wasps attack the cactus-feeding mealybug in sufficient numbers?
The confusion over how many mealybug species have been introduced – and where – severely hampered development of a program. (The mealybug has been introduced to control invasive cactus in Australia and South Africa. Most sources say it has been highly effective – prompting the initial concern when it appeared on Puerto Rico.) I have been unable to find any information about the status of the candidate biocontrol agents more recent than 2022.
Genetic Conservation
The USDA also partnered with the Naples (Florida) Botanical Garden to collect fruits and vegetative material for ex situ conservation. Rigorous phytosanitary procedures were followed to ensure the absence of the mealybug. Collections of fruits and vegetative material provided 1,298 cacti samples from 13 species, representing 1,173 maternal lines from 91 sites throughout Puerto Rico. A total of 90,720 seeds representing 8 species are banked at the NBG for long-term storage. Propagation of the vegetative material has 56% success, and plants are incorporated into the NBG’s living collections. (These figures include Opuntia cacti that are hosts of a second invasive insect, Cactoblastis cactorum.)
Genetic Concerns
Scientists now consider Hypogeococcus pungens (Hemiptera: Pseudococcidae) to be a species complex composed of at least five putative species. The species are separated in part by the plants they use as hosts. Two of the complex have apparently been introduced to Puerto Rico: H. pungens Hyp‑C feeds on cacti; H. pungens Hyp‑AP feeds on hosts in the Portulacaceae & Amaranthaceae. Both evolved from putative source populations in Brazil (Poveda-Martinez et al. 2022).
The two species H. pungens Hyp-C and Hyp-AP are currently separated on Puerto Rico by host preferences and climatic niches. They also occupy different geographic areas. Scientists fear that ongoing climate change could allow H. pungens Hyp-C to establish farther into the island’s interior and in a large area in the north. Such range expansion would end the geographic separation. Overlapping of the two species is likely to exacerbate the threat to Puerto Rico’s cacti. Most directly, it would complicate implementation of management strategies, especially biological control. Intermixing of the two species could also facilitate hybridization which might result in more vigorous attacks or a broadened host range. Hybridization is frequent in closely related species (Poveda-Martinez et al. 2022).
The Mealybug is Frequently Introduced
Mealybugs that feed on cacti and believed to be in the species Hypogeococcus pungens made multiple appearances in southern California between 2000 and 2018 – in gardens and in nurseries. Confusingly, CDFA reports interception of the mealybug on alternanthera and ludwigia plants shipped from Florida (CDFA 2018). I have no more recent data. The population in Florida was reported to be present in 16 counties in 2009); it might be the species that feeds on plants other than cacti (Poveda-Martinez et al. 2022). Other populations has been reported in the Dominican Republic (no date) (CDFA 2018); and in Hawai`i in 2005 (Hawaii Department of Agriculture new pest report). A mealybug that feeds on Amaranthaceae and Portulacaceae was detected in 2000 in San Juan, Puerto Rico (Poveda-Martinez et al. 2022).
In the absence of control measures, scientists expect H. pungens Hyp-C to continue decimating Puerto Rican cactus diversity and threaten other cactus rich ecosystems across the Caribbean islands, Central America and, potentially, North America (Poveda-Martinez et al. 2022).
saguaro and organ pipe cacti in Organ Pipe Cactus National Monument; photo by F.T. Campbell
North America has more than 500 columnar cactus species in the Cactoideae (Zimmerman et al. 2010). Some of these cacti are already endangered, e.g., several Pediocactus. Others are totems of the desert, e.g., the saguaro (Carnegiea gigantea) and organ pipe (Stenocereus thurberi) cacti. Picture The larger ones, particularly, play important ecological roles. It is not known how vulnerable individual species are to the mealybug (Golubov pers. comm. January 2011). In Mexico several mealybugs in the same genus are already present. The natural enemies of these mealybugs might be able to attack H. pungens Hyp-C if it invades the country (Zimmerman et al. 2010). Despite the well-founded concern, apparently no funds have been allocated by governments or conservation organizations to studying the vulnerability of these cacti to one or more mealybugs in the Hypogeococcus genus.
The most likely pathway by which the mealybug is spread is the trade in plants for planting (the horticultural trade) (Zimmerman et al. 2010). A decade ago APHIS reported intercepting mealybugs on cactus (primarily on roots) imported from Germany, Peru, and Puerto Rico. APHIS has also intercepted several other mealybugs in the same genus – on plants (including orchids and bromeliads as well as cacti) from Belize, Costa Rica, Ecuador, Guatemala, Honduras, Mexico, Panama, Peru, and Venezuela (USDA APHIS alert).
A decade ago NatureServe and IUCN found that 31% of Earth’s cactus species were threatened with extinction. They named overharvesting (often for the illegal horticultural trade) and destruction of habitat by smallholder livestock ranching and farming. Did not mention predation by introduced insects – although that is now manifest not only in the cactus mealybug but also the cactus moth.
Sources
Aguirre, M. G. Logarzo, S. Triapitsyn, H. Diaz-Soltero, S. Hight, O. Bruzzone. 2023? Effect of egg production dynamics on the functional response of parasitoids
California Plant Pest and Disease Report. 2005. Vol. 22 No. 1. Covering Period from July 2002 through July 2005.California Department of Food and Agriculture. 2018.California Pest Rating for Hypogeococcus pungens Granara de Willink | Harrisia cactus mealybug Hemiptera: Pseudococcidae Pest Rating: A California Pest Rating for Hypogeococcus pungens Granara de Willink | Harrisia cactus mealybug Hemiptera: Pseudococcidae Pest Rating: A
Poland, T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019), Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag.
Poveda-Martinez, D. N.A. Salinas, M. Belen Aguirre, A.F. Sanchez-Restrepo, S. Hight, H. Diaz-Soltero, G. Logarzo, and E. Hasson. 2022 Geonomic & ecol evidence shed light on the recent demographic history of two related invasive insects. Scientific Reports.
Segarra-Carmona, A.E., A. Ramirez-Lluch. No date. Hypogeococcus pungens (Hemiptera: Pseudococcidae): A new threat to biodiversity in fragile dry tropical forests.
Segarra-Carmona, A.E., A. Ramírez-Lluch, I. Cabrera-Asencio and A.N. Jiménez-López. 2010. First Report of a New Invasive Mealybug, the Harrisia Cactus mealybug Hypogeococcus pungens (Hemiptera: Pseudococcidae). J. Agrie. Univ. RR. 94(1-2):183-187 (2010)
Srivastava, M., P. Srivastava, R. Karan, A. Jeyaprakash, L. Whilby, E. Rohrig, A.C. Howe, S.D. Hight, and L. Varone. 2019. Molecular detection method developed to track the koinobiont larval parasitoid Apanteles opuntiarum (Hymenoptera: Braconidae) imported from Argentina to control Cactoblastis cactorum (Lepidoptera: Pyralidae). Florida Entomologist 102(2): 329-335.
Triapitsyn, Aguirre, Logarzo, Hight, Ciomperlik, Rugman-Jones, Rodriguez. 2018. Complex of primary and secondary parasitoids (Hymenoptera: Encyrtidae and Signiphoridae) of Hypogeococcus species. mealybugs (Hemiptera: Pseudococcidae) in the New World. Florida Entomologist Volume 101, No. 3 411
USDA Agriculture Research Service, Research Project: Biological Control of the Harrisia Cactus Mealybug, Hypogeococcus pungens (Hemiptera:pseudococcidae) in Puerto Rico Project Number: 0211-22000-006-10 Project Type: Reimbursable
Zimmermann, H.G., M.P.S. Cuen, M.C. Mandujano, and J. Golubov. 2010. The South American mealybug that threatens North American cacti. Cactus and Succulent Journal. 2010 Volume 82 Number 3
Prostanthera cuneata – member of a genus endemic to Australia. Photo by Leonora (Ellie) Enking via Flickr
In 2023 a global meeting of plant conservation experts convened by The Royal Botanic Gardens, Kew (U.K.) released the 5th edition of a report on the State of the World’s Plants and Fungi.
Associate Professor of Plant Ecology and Conservation Science Rachael Gallagher from Western Sydney University had led the global evaluation of conservation assessments for unique flora species. She is also the lead author of an article (2023; full citation at the end of this blog) evaluating how well countries around the world met their treaty obligation to assess the conservation status of endemic plant species native to their territories. The analysis identified 221,399 endemic plant species in a total of 173 countries. The treasure is not distributed evenly. Five countries harbor a third of the endemic plant species: in descending order, Brazil, Australia, China, Mexico, and South Africa. (The United States, including its islands, ranks 8th.)
On average, countries completed assessments of just 34% of their endemic species. New Zealand and here and South Africa shone: they assessed 87% of their unique species. China assessed 71%. One of the world’s poorest countries, Madagascar, evaluated 42% of its ~10,000 endemic plant species. Reminder: tiny Madagascar ranks 6th in the number of endemic plants. Australia – one of the richest countries– carried out the process for 39% — slightly more than the global average. Other countries that are stewards of numerous endemic plants were below the average: Brazil reviewed 29%, Mexico assessed only 24%.
Rachael Gallagher and her colleagues in the Australian Biodiversity Council were quite critical of Australia’s low level of performance. They called on their countrymen to do much more to prevent the decline and extinction of the country’s unique plant species. Australia, as party to the Convention on the Conservation of Biological Diversity, has a treaty obligation to prevent extinction of species which occur nowhere else. Remember, Australia’s flora and fauna rank extremely high on a scale of phylogenetic distinctness as an heir of the isolated continent of Gondwanaland.
Gallagher and colleagues concede that many endemic plant taxa in Australia have huge ranges — averaging 235,829 km2. But these vast expanses do not prevent sudden population crashes caused by calamities. They mention the megafires of 2019–2020 and – over the longer term – climate change. I think of the invasion by the rust fungus Austropuccinia psidii.
When we think about Australia, we wonder at the kangaroos and koalas. I assume Australians consider their unusual fauna to be iconic symbols of their country. Why are they not equally committed to their flora – 88% of their plant species are endemic. Do they suffer from the same “plant blindness” I have encountered in the United States? South Africa undertook an assessment of her endemic flora that concluded that a quarter of these species are threatened. Sixty percent of the country’s 20,000 plant species are endemic.
a protea in South Africa’s fynbos; photo by Michael Wingfield
[I have found no parallel analysis of America’s endemic plant species. Our nation’s rank of 8th in number of endemic species is explained by the highly unique floras of the islands, especially the Hawaiian archipelago. More than 95% of native species on the Islands are endemic. This includes 67% of the large trees still present in the forests (Potter et al. 2023).]
This study reflects the findings of the International Union for the Conservation of nature (IUCN)’s 2024 Red List of Threatened Species. A decade-long global project had found that at least 16,425 of the 47,282 tree species (38%) assessed are at risk of extinction. Trees accounted for over one quarter of species on the IUCN Red List. Tree species are at risk of extinction in 192 countries around the world.
Sources
Gallagher, R.V., S. P. Allen, R. Govaerts, M.C. Rivers, A.P. Allen, D.A. Keith, C. Merow, B. Maitner, N. Butt, T.D. Auld, B.J. Enquist, W.L. Eiserhardt, I.J. Wright, J.C.O. Mifsud, S. Espinosa-Ruiz, H. Possingham, V.M. Adams. 2023. Global shortfalls in threat assessments for endemic flora by country. Plants, People, Planet. DOI: 10.1002/ppp3.1036
Potter, K.M., C. Giardina, R.F. Hughes, S. Cordell, O. Kuegler, A. Koch, E. Yuen. 2023. How invaded are Hawaiian forests? Non-native understory tree dominance signals potential canopy replacement. Landsc Ecol 2023 https://doi.org/10.1007/s10980-023-01662-6
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
I last blogged about bioinvasions by earthworms in September 2023. My concern was that the USDA Animal and Plant Health Inspection Service (APHIS) was refusing to regulate worms’ introduction based on an inadequate risk assessment. I complained that APHIS’ principal concern was that introduced worms or the soil they were transported in might transport pathogens. APHIS’ website did not mention introduced earthworms’ well-established ability to disrupt the soil and possibly cause undesirable impacts on plant growth and diversity.
I also cited a study by USDA Forest Service scientists regarding interactions between long-term, continuous stressors, including earthworm invasions, and short-term gap-forming events. Reed, Bronson, et al. (2025) reported finding lower earthworm biomass and density in both deer exclosures and canopy gaps. They hypothesized that in canopy gaps higher deer browse pressure changed plant community and soil properties. These changes then affect soil-dwelling fauna like earthworms. They believed the higher worm densities in closed-canopy sites might be the result of greater volumes of nutrient-rich tree leaf litter which provides both shelter and food. The closed canopy sites also lacked recent soil disturbances. (Other authors cited in that blog describe varying impacts of deer browsing on plant communities and succession.)
A study I have not previously included in my blogs focused on invasions in the forest understory of the Katharine Ordway Natural History Study Area, which is in eastern Minnesota on the bluffs of the Mississippi River. The now dominant oaks are being replaced by a more mesic forest with a growing proportion of Ulmus americana, U. rubra, Acer negundo, Prunus serotina, Tilia americana, and Fraxinus pennsylvanica. (I note that three of these species are being suppressed by two introduced organisms, “Dutch” elm disease and emerald ash borer).
Davis et al. (2015) found that all common herb layer species – native and non-native – established in microsites favorable to plants in general. Under the forest’s mostly closed tree canopy, light levels played almost no role in determining where herbaceous understory plants grew. However, abundant earthworms and deer were deemed to be larger factors explaining most of the changes in native herbs’ abundance and distribution. The earthworms’ effect arose from changes to the soil and litter environment.
I have not found a more recent analysis of the Ordway forest understory, so I don’t know whether these trends continued. I also wonder whether the conclusions about herb species’ interactions would have differed if the non-native species studied were those that completely dominate the ground level, such as Ficaria instead of Alliaria petiolata; or a dense shrub layer, e.g., Rosa multiflora.
A new analysis provides a large-scale description of worm invasions. McCay et al. (2026) report that about 300 species of earthworms inhabit North America, of which about 70 are non-native. They sought to determine the relative importance of Pleistocene glaciation in explaining earthworms’ present assemblages. Their study sampled 94 sites across 13 states of the eastern and central U.S. over a twelve year period (2013 – 2025). Seventy-four of the sites were located in previously-glaciated locations, 20 sites in non-glaciated places.
The study confirmed the presence of 16 species; only three were considered to be native to North America (Aporrectodea rubidus, Eisenoides carolinensis and E. lonnbergi). Minimum species richness at individual sites ranged from zero to eight species. Species richness was markedly greater in the glaciated regions of the Midwest and Northeast. By both dry biomass and numbers, European lumbricids dominated throughout. Lumbricus species constituted 45% by number and 69% by biomass across all sites. Their dominance was highest in the Midwest Glaciated region.
Native earthworms were rare, constituting only 2 or 3% at those sites where present. Invasion by Asian pheretimoid megascolecid (“jumping worm”) species has just begun; they were present at only 10% of study sites.
Earthworms – apparently both native and non-native – were most abundant and diverse in the Midwest Glaciated region (Ohio, Michigan, Minnesota, and Wisconsin), where non-indigenous earthworm invasion is more recent than in the Northeastern Glaciated Region (Massachusetts and New York). Earthworms were least common and diverse in areas south of the terminus of the Pleistocene glaciation. McCay et al. (2026) speculated that development of forests in northern North America over thousands of years with few or no earthworms might have led to accumulations of organic matter in forms useful to earthworms (“banking” of soil organic matter), which supported a pulse of earthworm proliferation following colonization.
Some other factors play a role. Earthworm presence – measured by biomass – and diversity increased with higher soil pH. Many earthworm species are known to be sensitive to soil acidity. Epigeic earthworms (those that feed on leaf litter) are often more tolerant of acidic conditions than endogeic earthworms (which live in and consume soil).
The study did not measure co-occurring variables such as calcium abundance, buffering capacity, or exchangeable aluminum. All might more directly affect worms’ physiology than pH per se.
Soil structure also matters. Soil-feeding species might be disadvantaged by sandy and gravelly soils, which do not support persistent burrow formation and are often nutrient deficient. On the other hand, they thrive better in clay soils.
While each study participant established at least one site in a relatively undisturbed habitat, e.g., a mature forest, the majority of sites tended to be within a disturbed landscape, less than .8 km from a road.
Earthworm abundance and activity vary seasonally but only 4 sites were observed over more than one season. Across all sites, there was greater species richness in autumn and spring than summer. McCay et al. (2026) think that this might have been because many earthworms aestivate during dry weather, so they are less detectable during summer although they are still present.
Distribution of native North American earthworms suggests there was some natural colonization of previously glaciated landscapes before European people arrived.
McCay et al. (2026) also did not document land use history at the sampling sites, although that can profoundly affect worms and vertical distribution of carbon in the soil. Nor did their habitat classification system [deciduous and coniferous forests, mowed and unmowed grasslands] describe the vegetation in sufficient detail to determine whether food quality might have been a factor. It is recognized that litter varies in C:N ratios, lignin content, and palatability.
McCay et al. (2026) note an interesting pattern at the global scale: Australia, New Zealand, South Africa and North America (U.S. and Canada) have been invaded by the same suite of invasive European earthworm species. One common factor, not addressed by the scientists, is that all were settled by British colonists. Instead, they focus on ecological factors. They suggests that the species’ niche requirements are broad and well-matched to temperate forest soils worldwide, and that climate and soil conditions, rather than biotic resistance from native communities, are primary factors limiting their distribution and abundance.
Another factor supporting this hyptothesis, in their view, is that the composition of earthworm communities was relatively similar above and below the southern glacial limit in North America. Here, European earthworms colonized not just the relatively “open” soils of previously glaciated north but also non-glaciated areas farther south – assisted by human introductions and creation of disturbed habitats. They suggest that the novel species might have benefitted from reduced competition because the European species had functional differences from resident native species.
The current rapid expansion of “jumping” earthworms in North America is a fundamentally different invasion wave than the centuries-old invasion by European lumbricid species. The Asian pheretimoid species reproduce parthenogenetically and have shorter generation times. They achieve high population densities at the soil surface, process organic matter rapidly, and create a loose casting layer, potentially creating different soil conditions in ways not true after invasion by European earthworms. Although it is too early to know the outcome, McCay et al. (2026) note emerging evidence that the Asian pheretimoids might outcompete European earthworms through more flexible resource exploitation, preemptive foraging during the late summer and early fall, and tolerance of environmental stress.
Effects on atmospheric carbon levels
McCay et al. (2026) point out that the introduced earthworms are disrupting the significant pool of accumulated organic matter in previously glaciated soils. As earthworms move in, they facilitate more rapidly cycling but also translocate carbon into deeper soil strata. They conclude that while earthworms initially accelerate carbon loss through increased decomposition and respiration, their long-term effects might protect soil carbon storage. Earthworm casts and burrows can create microsites with different oxygen and moisture conditions that might promote carbon stabilization through different mechanisms than the original forest floor.
SOURCE
Davis, M.A., M.D. Anderson, L. Bock-Brownstein, A. Staudenmaier, M. Suliteanu, A. Wareham and J.J. Dosch. 2015. Little evidence of native and NIS species influencing one another’s abundance and distribution in the herb layer of an oak woodland. Journal of Vegetation Science · June 2015
DOI: 10.1111/jvs.12302
McCay, T.S., Anderson, L.J., C.P. Bloch, A.E. Cahill, S.L. Cooke, B.J. Dolan, K.M. Flinn, D. Garneau, N.J. Hains, K. Hopfensperger, M.Beth Kolozsvary, C. Mankiewicz, S.E. Scanga, J.L. Schafer, E. Schwartzberg, D.A. Scott, K.L. Shea, J. Simmon, J.N. Styrsky. 2026. Earthworm assemblages in the Eastern and Midwestern United States and the legacy of glaciation. Biol Invasions (2026) 28:91 https://doi.org/10.1007/s10530-026-03798-3
Reed, S.P., D.R. Bronson, J.A. Forrester, L.M. Prudent, A.M. Yang, A.M. Yantes, P.B. Reich, and L.E. Frelich. 2023. Linked disturbance in the temperate forest: Earthworms, deer, and canopy gaps
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
eastern (Canadian) hemlocks in Nova Scotia killed by hemlock woolly adelgid; photo by Celia Boone, NSDLF
Two recent studies show that climate change is driving changes to pest ranges. These pose a serious threat to forests of eastern Canada.
A study by Aubin et al. (2026; full citation at end of blog) assessed the risk that climate change would exacerbate the impacts of 14 non-native forest insect pests The 14 insect species were selected from an original group of 76 species using the following criteria:
1) They have the potential to cause mature tree mortality to at least one of the 37 tree species most abundant in Canada; and
2) They have been detected in Canada recently or have established populations actively spreading through the Canadian landscape.
Major Findings
Twenty-four tree species are affected currently by at least one of the 14 non-native insect pests (excluding Asian longhorned beetle; see below). (I list the tree and insect species below.) Four of them are already considered globally endangered or threatened due to invasive pests: black and white ash (Fraxinus nigra and F. americana), whitebark pine (Pinus albicaulis) and eastern hemlock (Tsuga canadensis). Another ash species (blue ash; Fraxinus quadrangulata) and another pine (limber pine; Pinus flexilis) are considered rare or threatened species in Canada.
brown spruce longhorned beetle; photo by Udo Schmidt via WikiMedia
Of the 24 species at risk, black spruce (Picea mariana) is most exposed because it grows in areas where eastern spruce gall adelgid and brown spruce longhorned beetle (Tetropium fuscum) are established. Expansion of these two pests could potentially reach 75% of black spruce biomass in Canada. However, Aubin et al. (2026) expect tree mortality to be limited because these insects target trees that are already stressed or weakened. Of course, the changing climate might increase the trees’ susceptibility. Thirty percent of the black spruce’s range is projected to be outside its current climatic niche by 2040.
The 24 tree species currently affected by at least one of the 14 non-native insects (excluding ALB) collectively constitute 3.2 billion tons of tree biomass. The impact is projected to increase more than four-fold — to 13.6 B tons of biomass — within two decades. Reinvasion by the Asian longhorned beetle would put at risk six additional tree species in the genera Acer, Betula, and Populus. Their jeopardy would add another 3.1 billion tons of live tree biomass to the “at risk” category.
Aubin et al. (2026) note that affected trees might remain alive but moribund, that is, lose their functional role within the ecosystem. for several years before finally dying. Therefore their analysis might underestimate pests’ impact on the forest. Their example is American beech – as weakened by beech bark disease. A reminder: beech bark disease causes widespread death of mature beech – opening the canopy and eliminating such wildlife-supporting aspects as nesting cavities and abundant crops of nuts. Vigorous root sprouting results in dense stands of young beech, crowding out other species.
Hotspots of greatest vulnerability
The analysis identified two hotspots of greatest vulnerability: north-central British Columbia near the border with Alberta, and along the St. Lawrence Seaway near the border with the United States. The pest pressures differ.
The threat to the British Columbia hotspot comes from expansion of mortality in dense pine forests caused by the native mountain pine beetle (Dendroctonus ponderosae). Aubin et al. (2026) describe a two-pronged impact from the changing climate: trees lose vigor because they become maladapted to the new growing conditions (temperature and drought); while the beetle increases the frequency of outbreak due to reduced overwintering mortality.
Along the St. Lawrence Seaway (southern Ontario and Quebec) the threat comes from a suite of non-native insects, including emerald ash borer, hemlock woolly adelgid, and beech scale. While emerald ash borer and beech scale were introduced directly to Canada by international trade, hemlock woolly adelgid spread across the border from the U.S. Further northward expansion of all three is projected under both low and high emission climate change scenarios.
beech bark disease in New Hampshire; photo by Eli Sagor via Flickr
Most alarming is that some regions in eastern Canada are vulnerable to invasion by all 14 insect species. Two additional pests loom: Aubin et al. (2026) fear northward expansion might reintroduce the Asian longhorned beetle or introduce the southern pine beetle (Dendroctonus frontalis). The latter has been expanding northward in the U.S.
Pests often move across the Canada-U.S. border. In addition to the five pests mentioned above, spongy moth, and two pathogens, the beech leaf disease nematode, and the oak wilt fungus, have spread from the U.S. into neighboring parts of Canada. The woodwasp Sirex noctilliowas probably introduced simultaneously to both countries. Winter moth and beech bark disease spread from Canada to the U.S. I worry that the brown spruce longhorned beetle might do the same.
Asian longhorned beetle: can this disaster be averted?
As noted above, the Canadians are alarmed by the prospect that the Asian longhorned beetle might be reintroduced – either by spread from extant populations in the United States or directly on imports from China. Aubin et al. (2026) note that the two earlier – successful! — eradication programs were expensive, costing an estimated CND$35.5 million. This expenditure is dwarfed by the costs estimated to arise from an unmanaged invasion: CDN$431 million annually in timber products and CDN$358 million annually in edible maple products. There would also be enormous ecological impacts, including threats to an additional 3,08 metric tonnes of tree biomass comprising ~24.96 tons of CO2 equivalent.
Canada’s central boreal forest is at lower risk both in terms of exposed tree biomass and number of invasive insect species present. The reasons are not understood. Aubin et al. (2026) suggest that the boreal ecosystem is more resistant to invasion due to a combination of environmental barriers and native natural enemies. For example, the introduced woodwasp Sirex noctilio did not cause widespread pine mortality in the region, probably due to antagonistic interactions with other subcortical species.
Although it does not fit this definition, Aubin et al. (2026) also evaluated the Asian longhorned beetle (ALB Anoplophora glabripennis), because of its huge impact if it is reintroduced to Canada (see above).
One of the focal groups, beech scale, Cryptococcus fagisuga, differs from the others because it is a vector of a tree-killing fungal pathogen (Neonectria coccinea); the scale itself does not cause notable harm.
Aubin et al. (2026) acknowledge that additional species represent a possible threat to Canadian forests. Therefore their study does not represent the total risk posed by all potential invasive insects in Canada, but provides a snapshot of selected, current vulnerabilities.
The 37 most abundant tree species in Canada together represent 88% of total mature forest tree biomass in Canada. They include 17 conifers and 7 deciduous trees:
Abies amabilis, Ab. balsamea, Ab. lasiocarpa; Acer rubrum, Ac. saccharinum, Ac. saccharum; Alnus rubra; Betula alleghaniensis, B. papyrifera; Callitropsis nootkatensis; Fagus grandifolia; Fraxinus american, F. nigra; Larix laricina, L. occidentalis; Picea engelmanii, P. glauca, P. mariana, P. rubens, P. sitchensis; Pinus albicaulis, P. banksiana, P. contorta, P. ponderosa, P. resinosa, P. strobus; Populus balsamifera, P. grandidentata, P. tremuloides; Pseudotsuga menziesii; Quercus rubra; Thuja occidentalis, T. plicata; Tilia Americana; Tsuga canadensis, T. heterophylla, T. mertensiana
The 14 insects collectively have 63 host tree species in Canada. The pine genus is susceptible to the largest number of pests. Genera found to be not vulnerable to any of the 14 insects are Acer, Callitropsis, Populus, Pseudotsuga, Tilia, and Thuja. The proportion of total exposed tree biomass in Canada varied by species, from 8% of Jack pine (Pinus banksiana) to 95% for red spruce (Picea rubens).
red spruce in West Virginia; photo by Famartin via WikiMedia
Although most of the 14 insect species are projected to benefit from larger areas of suitable climate in Canada over the next 20 years, there are interesting exceptions: European oak borer (Agrilus sulcicollis), brown spruce longhorned beetle, and winter moth (Operophtera brumata). Distributions of the host tree species are projected to change insignificantly over the 20 years covered by the study.
With northward expansion of suitable climates for 12 of the 14 species, large areas of the boreal forest will be exposed to potential invasion. The entire Canadian distribution of 13 of the 37 dominant tree species might be at risk: three Abies, two Betula, both Fraxinus, Picea mariana, Pinus contorta, Picea glauca, Pinus banksiana. Aubin et al. (2026) mention specifically Engelmann spruce (Picea engelmanii) and American beech (Fagus grandifolia).
Other Factors
The analysis did not consider possible alterations of the insects’ life history traits other than potential expansion of their distributions. Warmer temperatures can cause changes in voltinism, diapause periods, development rates, reproduction, and population growth; cumulatively, these changes might alter their invasion dynamics. The mountain pine beetle is an example. Populations have experienced outbreaks more frequently, so increasing the species’ invasion threat and severity. On the other hand, shifts in temperature and precipitation could decouple the phenology of trees and their associated pests, reducing insect survival. Finally, complex changes in tree tissue and their secondary defensive metabolites (see below) also could alter interactions between non-native insects and their new hosts – possibly exacerbating or mitigating the herbivores’ impacts.
Aubin et al. (2026) remind us that loss of a dominant species might lead to reorganization of forest composition and structure. They expect the impacts to be particularly critical in stands with low tree diversity, such as the pure jack pine stands in the eastern boreal forest. Loss of a foundation species might also profoundly disrupt ecosystem functions, carbon budgets, wildlife habitats and stand productivity. They cite cascading effect on aquatic invertebrate communities and invasions by non-native plant species following widespread death of ash trees caused by the emerald ash borer. Finally, death of some species reduces functional redundancy within tree communities, and a shrinking pool of viable native replacement species. The widespread planting of ash trees in urban areas after the demise of most elms is such a case.
American elms in Jeanne d’arc Park, Quebec; photo by Cephus via Wikimedia
The possibility that the tree hosts might increase production of defensive metabolites was corroborated by Mike Aucott in a different context. Dr. Aucott is retired from the New Jersey Department of Environmental Protection. He authored a guest blog in December 2022, in which he discussed changes in plant chemistry brought on by the 50% increase in atmospheric CO2 levels over the last century. By happenstance, Dr. Aucott engaged in an exchange of letters in Science (2/26/26), in which he reiterated the likelihood that plants, “fertilized” by access to this nutrient, might be better able to fend off insect attacks. (See the “Sources” section for references to additional information on this phenomenon.)
Compounding Threat: The Spruce Budworm in Eastern Canada
The spruce trees of eastern Canada face another pest threat: the native spruce budworm (SBW, Choristoneura fumiferana). It is already a major defoliator in North American boreal forests.
Boulanger et al. (2025; full reference at end of this blog) documented pronounced changes in the moth’s range, especially in the East. Over the past 60 years, suitable climate conditions for the budworm have expanded northward. On the other hand, winter mortality has increased in southern parts of its range due to warmer temps. Overall, the total area highly suitable for population growth remained virtually the same. Still, the budworm’s earlier activation might exacerbate its impact on the previously less vulnerable black spruce, Picea mariana. If so, this might fuel further increased population growth rates northward.
black spruce; photo by Laval University via WikiMedia
Like Aubin et al. (2026), Boulanger et al. (2025) found that Canadian forests in the east and Atlantic regions are likely to experience greater impacts on tree growth than are forests in western and central regions. The host most vulnerable to SBW, balsam fir (Abies balsamea), is a dominant or codominant species in the East and Atlantic regions. The fir is sparsely distributed in those areas of central and western Canada where the climate is becoming highly suitable for the insect. The frequent wildfires promote growth of young pioneer tree species, e.g., jack pine and trembling aspen, that do not support SBW.
At the most general level Boulanger et al. (2025) suggest that climate change might have already surpassed impacts of land use change on spruce budworm dynamics. Climate change puts additive and synergistic pressures on insects, which are already more sensitive than trees to climatic factors and able to adapt more quickly. As a result, climate change is becoming the most significant driver of recent declines in insect abundance and shifts in community structure, development, dispersal patterns, and phenology. (Again, see Aucott, above, for an alternative explanation.)
Boulanger et al. (2025) mention but do not discuss possible impacts of climate change and a shift in SBW distribution and tree hosts on a third trophic level, i.e., natural enemies. They note that many factors – not just climate suitability — influence trophic interactions. Another complication is that most SBW parasitoids require alternate hosts.
Boulanger et al. (2025) join others in urging forest managers to quickly adapt their management strategies to the novel climate-induced threats. They call for a proactive and integrated approach in forest management. Ecological research will be crucial to mitigate the compounded effects of climate change and to preserve the integrity & sustainability of forest ecosystems.
SOURCES
Aubin, I., A. Roe, B. Marquis, L. Boisvert-Marsh, J. Pedlar,S. Erni, B. Hamel, G. Lawrence, D. McKenney, T. Scarr. 2026. Vulnerability of Canadian forests to invasive insects under climate change. Accepted by the Canadian Journal of Forest Research.
Boulanger, Y., A. Desaint, V. Martel, M. Marchand, S. Massoda Tonye, R. Saint-Amant, et al. (2025) Recent climate change strongly impacted the population dynamic of a North American insect pest species. PLOS Clim 4(2): e0000488. https://doi.org/10.1371/journal. pclm.0000488
See also an article describing declining nutrient value of food crops in response to increased atmospheric C02 levels: https://www.washingtonpost.com/climate-environment/interactive/2026/carbon-pollution-diluting-key-nutrients-food/
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The Wildlife Society’s Invasive Species and the Forestry and Wildlife Working Groups are cosponsoring a workshop for the Lingering Hemlock Project. The workshop will be on Tuesday 28 April at 2:00 PM EDT.
The Lingering Hemlock Project is a subset of The Nature Conservancy’s “Tree Species in Peril” program. The project aims to locate and selectively breed eastern hemlocks with genetic resistance to the hemlock woolly adelgid (HWA).
Olivia Hall from the North Carolina Hemlock Restoration Initiative will share more about how natural areas can participate in the Lingering Hemlock Project. In the southeastern US, project partners can locate and record data on hemlocks that remain healthy despite HWA infestations. In the northeastern US, project partners can establish hemlock plots and monitor their health and decline annually.
Go here to learn more about the project & webinar, and find the link to join.
I have blogged about HWA often – although there is no simple method for finding the earlier blogs. In 2025 I posted 3 blogs – in March and one in August. If you need a reminder about HWA, visit TNC’s “don’t move firewood” website here.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Amynthas agrestis; photo by John Abrams via Wikimedia
The University of Minnesota is seeking to learn the extent and impact of invasive Asian jumping worms (Amynthas spp.). Scientists fear that jumping worms will remove the naturally deep litter layer and create extremely loose soils that cannot be held by plant roots. These changes will expose soil on the state’s hillsides to erosion by human footsteps, rainfall, and water runoff. They worry about the future sustainability of forested hills in Minnesota.
The research project began in January 2024; it is funded at $430,000. The research seeks to answer the following questions:
What is the magnitude and rate at which jumping worms accelerate soil erosion in forested hillslopes in Minnesota?
What are the mechanisms of soil erosion by jumping worms in hardwood forests?
What is the spatial extent of jumping worms in forested hillslopes?
What species of native plants are capable of holding soils against jumping worms and could act as erosion-prevention?
What management practices could help to reduce soil erosion induced by jumping worms in forested hillslopes?
The scientists are asking volunteers to actively look for jumping worms in the forests of southeastern Minnesota and report them to EDDMapS.
Boundary Water Canoe Area; photo by Chad Fennell via Wikimedia
The scientists remind us that invasive European earthworms have already infested nearly all of the state’s forests, even in the remote Boundary Waters Wilderness. Forest soils and understory vegetation transformed, and invasive earthworm impacts are cascading through ecological and socio-economic processes. They expect the state to become divided into two distinct areas, each dominated by a different invasive earthworm group.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Lindera benzoin; photo by Fritzflohrreynolds via Wikimedia
Scientists in Ohio and other states are trying to determine what is causing dieback of the native shrub northern or common spicebush (Lindera benzoin). The shrub occurs throughout much of the eastern United States and neighboring parts of Ontario, Canada.
In May 2024, Ohio State forest entomologist Kayla Perry and Cleveland Metroparks Natural Resources Area Manager Josh Philipps observed red thrips and dieback symptoms on spicebush in the reserve. The website includes many photographs of the damage. [Interesting note: beech leaf disease was also first detected in Cleveland Metroparks.] Later in the season similar symptoms were detected at Holden Arboretum and in other units of Cleveland Metroparks properties across three Ohio counties, Cuyahoga, Lake, and Medina. Constance Hausman, Senior Conservation Science Manager with Cleveland Metroparks, reported seeing dieback on spicebush at other reserves in Lake, Ashtabula, and Geauga Counties, suggesting the problem is likely more widespread.
At some locations, large populations of thrips were found on a few plants where they were congregating on the underside of the leaves. While some of the plants show twig dieback, other plants wilt. Some had chlorotic leaves with a mottled pattern. Other symptoms included black necrosis of the petioles and some vein spotting. Similar dieback and viral symptoms observed at other parks within the region. At an arboretum in Wooster, symptoms were seen but no thrips were observed on those plants. As of December 2024, there was no evidence of a connection between the dieback symptoms and the presence of the thrips.
Ohio State entomologists identified the thips as nymphs belonging to the suborder Tubilifera or tube-tailed thrips. National Identification Services (NIS) confirmed the identification as Pseudophilothrips in the family Phlaeothripidae. However, a species determination could not be made due to the limited genetic database of Thysanoptera in GenBank or BOLD.
Some of the symptoms are often associated with a viral infection. Examination of a few twigs found no signs or symptoms of black twig borer (Xylosandrus compactus). Culture plates showed the growth of Colletotrichum sp., a well-known pathogen capable of infecting a wide range of host plants and causing various host-specific symptoms — often referred as anthracnose. The C. Wayne Ellett Plant Pest Diagnostic Clinic did not observe the characteristic vascular streaking associated with vascular streak dieback, a fungal disease caused by Ceratobasidium theobromae. (Spicebush with these symptoms were detected on 30 years ago at two locations in southwestern Ohio, but the cause was not determined.)
Scientists at USDA-ARS were testing symptomatic samples for another possible agent, the recently discovered Emaravirus. The December 2024 article said results were pending. However, at the February 2026 USDA Annapolis, participants reported no breakthroughs.
As of late 2024, symptomatic plants had been detected in Kentucky, Missouri, and Virginia, as well as Northeast Ohio. I believe Maryland has also found symptomatic spicebushes.
Lindera species are also threatened by laurel wilt disease, which is spreading north.
The three spicebush species are hosts for several native swallowtail butterfly species and other pollinators, including spicebush swallowtail (Papilio troilus), eastern tiger swallowtail (Papilio glaucus), and promethea silkmoth / spicebush moth (Callosamia promethea). I believe the shrubs support largely the larvae of these species. The grubs of the sassafras borer (Oberea ruficollis) will bore into the shrub’s branches and roots.
Spicebush is one of the first shrubs to bloom in Pennsylvania forests. I have found no information concerning the plants’ importance to early season pollinators other than the butterflies.
grey catbird; photo by Wilfred Hdez via Flickr
The red, shiny, elliptical fruits with a single seed (drupes) are nearly 50% fat. They become ripe in the fall, so a great fuel source for fall migrants and over-wintering resident birds, including wood thrush (Hylocichla mustelina), veery (Catharus fuscescens), northern bobwhite (Colinus virginianus), and gray catbird (Dumetella carolinensis). Eastern cottontail (Sylvilagus floridanus) and other small mammals might feed on the leaves, twigs, and berries. Other sources also mention deer. However, the spicy, sweet scent produced by the stems and foliage might deter some animals.
Spicebush leaves and berries can be used as a spice when cooking. Native Americans protected these plants used the plants to treat colds, coughs, and dermatological and respiratory ills.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The Trump Administration proposes (again!) to end all funding for USFS Research and State, Private, and Tribal Forestry programs. The budget document claims that these cuts are necessary “to ensure fiscal responsibility w/ taxpayer dollars & appropriate alignment of resources w/ USFS’s responsibility to appropriately steward National Forest System lands.” Ending the SP&T programs is justified as “better balance[ing] the appropriate roles of federal & State governments. … and [restoring] federalism …] The document claims that the federal component of Forest Health Management [currently receiving $16 million] duplicates programs managed by the National Forest System; yet the actions listed under this second budget category all relate to water management, not insects or pathogens. The document says states should manage pests on non-federal lands [currently receives $42 million]. I think this approach ignores the need for coordinated management for each of hundreds of pest species, from detection to eradication or development of host resistance. Eliminating the Research program will deprive all forest managers of a scientific foundation for management efforts.
The Trump Administration’s proposed budget would hold funding for key APHIS programs steady. This is great news compared to the extreme cuts proposed for the Forest Service. The budget document says that it is essential to continue APHIS programs success; any stoppages or reductions would potentially cause catastrophic consequences for environmental health. Contrary to this statement, holding funding steady actually results in cuts due to continuing introductions of new pests and inflation.
Item
2024 Actual
2025 Actual
2026 Estimated
2027 Estimated
Field Crop & Rangeland Ecosystems Pests (incl cogongrass)……….
12,000
12,000
11,000
9,026
Pest Detection ………………………………………………..
29,000
29,000
29,000
29,000
Plant Protection Methods Development ………………….
21,500
21,500
21,500
21,500
Specialty Crop Pests …………………………………………
215,000
215,000
214,000
217,339
Tree & Wood Pests …………………………………………..
59,000
59,000
58,650
58,650
Subtotal, Plant Health …………………………………….
387,500
387,500
385,150
386,515
USDA Forest Service
Two USFS programs w/ vital roles in protecting resilience of the Nation’s forests in the face of invasions by non-native pests and plants: R&D program and FHM within SPT division
The many economic & ecological benefits from our forests are under growing threats from a variety of disturbances, ranging from fires & hurricanes to non-native pests. ~ 60% of forests owned by non-feds; USFS must address threats to forests outside NFS to achieve its mission of sustaining “health, diversity, & productivity of the nation’s forests & grasslands to meet the needs of present & future generations.”
Research & Development
The Continuing Resolution for FY26 funded Research at $308 million for the year. Ask Congress to maintain this level. + increase research on invasive species from the current level of 1% to 5%.
The area of our forests & woodlands that is threatened by alien pests is similar to that attributed to fire or western bark beetles. More than 41% of forest biomass in the “lower 48” states is at risk to established non-native pests.[1] If able, add reference to pests on Hawai`i or Caribbean islands. Since additional introductions almost guaranteed, even greater proportion of US’ forest resources at risk in future. If possible, name example, e.g., Phytophthora austrocedri.Forest managers cannot counter these threats without understanding how these P&P kill trees & what actions are effective counter measures. This knowledge is obtained by research.
At least 53 tree species in forests across America are already under attack by non-native pests and pathogens. Yet as of FY23, Research stations spent just 1% of appropriation studying a few of the dozens of NIS pests. Funding for alien pests has decreased 70% since FY2010 even as new pests enter our forests. This inadequate research effort means USFS cannot develop effective programs to prevent, suppress, & eradicate the majority of alien pests. One crucial strategy suffers particularly = efforts to breed trees able to thrive despite NIS pests. R&D currently supports only a few such projects.
Forest Health Management: Supporting the Full Continuum of Pest Management
The Continuing Resolution for FY26 funded State, Private, and Tribal forests program at $310.6 million. I have not found specifics for the FHM program. This was an increase over the $281 million level in FY25.
Non-native pests and pathogens arrive as contaminants or hitchhikers on imported goods, especially on wood packaging and plants. These imports usually arrive in cities or suburbs, so the pests establish there first. They immediately cause enormous damage to urban forests, forcing local governments and property owners to absorb high tree removal costs. They then spread to rural forests, including National forests. Examples include hemlock woolly adelgid, emerald ash borer, invasive shot hole borers, goldspotted oak borer, sudden oak death, and beech leaf disease.
The most effective approach is to kill the pests where they first appear – usually in those urban or semi-rural forests. This response is led by FHM Cooperative Lands subprogram. We urge maintain funding for this subprogram at the FY26 level (possibly $42 million) so that the agency’s experts can continue to assist the states and other partners in countering these pests. As these pests spread to rural areas – including to National forests, National parks, and other public lands, responsibility for their management involves FHM Federal Lands subprogram. So much maintain funding for this subprogram at FY26 levels.
A recent analysis[2] determined that the natural resource values of 92 National parks are threatened by forest pests. Western parks are threatened primarily by outbreaks of the native mountain pine beetle (Dendroctonus ponderosae). Those in the East face threats from more than a dozen species of non-native pests, including hemlock woolly adelgid, emerald ash borer, spongy moth, laurel wilt, and – most recently – beech leaf disease.
Again, combatting these pests requires understanding their life histories & traits – understanding gained through the research program mentioned above.
Funding reductions over the past decade have already shrunk the number of FHM projects & areas treated each year. While 53 tree species are threatened, only four [eastern oaks, loblolly & ponderosa pines, & hemlocks] are targeted by 95% of projects. To counter the threats to 50 additional tree taxa, FHM needs additional resources.[3]
Investing in urban forestry is key to addressing both parties’ priorities & advancing flexible & cost-effective solutions to a wide range of issues impacting American communities, businesses, & families. The USFS SPT division’s Urban & Community Forestry Program efficiently distributes funds to shovel-ready projects for improving communities by maintaining a healthy tree canopy. Federal “seed” money provides resources necessary to initiate & stabilize these local programs.
A surprisingly high proportion of the (inadequate) funding for breeding trees to mitigate the damage caused by non-native pests is from FHM or the NFS, rather than R&D. These programs should receive substantial increases. The model program is the Dorena Genetic Resource Center. It provides decades-long commitment, skilled staff, necessary facilities; these result in breeding successes, i.e., western white pines and Port-Orford cedar.
Invasive Plants
Invasions of forests by non-native plant species erode forest productivity & provision of the full range of ecosystem services, hinder forest uses, degrade biodiversity & habitat, and impose substantial financial costs. A recent analysis[4] documents that this threat is growing: the number of FIA inventory plots containing invasive plant species rose in 58.9% of surveyed counties. Furthermore, in 73.2% of the counties the plots experienced an increase in species richness of invading plants. Increases occurred in all regions, but were greater in the East: from 46% to 52.3%. In the Rocky Mountains, the proportion of invaded plots rose from 6% to 11%. In Hawai`i, this proportion grew from 70% to 83.2%. Again, USFS Research and FHM programs, working together, are key to making progress in countering these bioinvasions.
[1] Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
[2] Michalak, J.L., C.E. Littlefield, J.E. Gross, T.G. Mozelewski, J.J. Lawler. 2026. Relative Vulnerability of US National Parks to Cumulative & Transformational Climate Impacts. Conservation Letters, 2026 Vol 19, Issue 1; 19:e70020
[3] Coleman, T.W, A.D. Graves, B.W. Oblinger, R.W. Flowers, J.J. Jacobs, B.D. Moltzan, S.S. Stephens, R.J. Rabaglia. 2023. Evaluating a decade (2011–2020) of integrated forest pest management in the United States. Journal of Integrated Pest Management, (2023) 14(1): 23; 1–17
[4] Potter, K.M., B.V. Iannone III, K.H. Riitters, Q. Guo, K. Pandit, C.M. Oswalt. 2026. US Forests are Increasingly Invaded by Problematic NIS Plants. Forest Ecology & Management 599 (2026) 123281
USDA Animal and Plant Health Inspection Service
APHIS is responsible for preventing intro and spread of pests and invasive plants that harm agric, including forests. APHIS policy guides port inspections carried out by the DHS CBP. APHIS inspects imported live plants.
Introductions of pests and pathogens have continued to occur. APHIS funding has remained steady – which means it is not growing to match the rising threat. At minimum, maintain current levels.
FY2025 enacted FY26 House FY26 Senate
APHIS total $1,148 $1,146 $1,168
Plant health subtotal $387.5 $388.6
Agric. quarantine $35.5 $35.5 $35.5
Field crop and rangeland $12 $11 $11.5
Pest detection $29 $28.5 $29
Methods development $21.5 $21.5 $21.5
Specialty crops $206.5 $216.3 $208.5
Tree and wood pests $59 $59 $58.6
Emergency preparedness and response* $44.5 $44.5 $44.3
* this fund is apparently for both animal and plant emergencies
Rationale
Already introduced pests threaten the many forest products and services benefitting all Americans. Just 15 of the worst pests threaten 41% of forest biomass in the “lower 48” states – comparable to fire.[1] A significant proportion of the resulting costs are imposed on municipal governments and homeowners. Fifteen years ago, it was estimated[2] that the municipal governments were spending more than $1B / year, primarily on removing and replacing trees on public property killed by these non-native pests. Homeowners faced costs of $1B plus loss of another $1.5B in property value. A more recent study estimated that cities will have to spend $30M per year to remove and replace ~ 1.4M street trees by 2050. Additional trees in parks and on homeowners’ properties also die.[3]
A new pattern has appeared in recent years: more newly-introduced pests are being detected in the Pacific Coast states rather than in the East and Midwest. Two southern California counties are projected to pay $150M – $1B[4] to remove and replace trees killed by invasive shot hole borers. The emerald ash borer threatens 9,000 ash on the streets of Portland, Oregon and millions more in parks and the forested wetlands of Willamette Valley, including in Ankeny National Wildlife Refuge. The Mediterranean oak borer has already killed thousands of oak trees in the San Francisco Bay area; it also threatens urban forests and valued oak savannahs in Oregon.
Additional introductions of highly damaging wood-borers are likely because we continue to receive inadequately treated crates, pallets, and other forms of packaging made of wood. For 20 years, all countries shipping goods to North America must treat their wooden packaging per prescribed protocols. To address this risk, we urge a modest $1M increase in APHIS’ “Tree and Wood Pest” account. We also suggest that the Subcommittee inquire of APHIS what steps it will take to improve compliance with the treatment requirement. You should focus your inquiry on China; wood packaging from this country is three times more likely to harbor a tree-killing pest than the global average.[5]
Other pests—especially plant diseases and sap sucking insects—enter on imported plants. Pathogens introduced recently via this pathway include rapid ohia death in Hawai`i (threatening the species that constitutes 80% of the Islands’ forest biomass) and beech leaf disease (thin a dozen years has spread across much of the East).
All assessments of APHIS’ plant import programs’ effectiveness use data from 2009; at that time, plant imports were more than 100 times more likely to transport pests than was wood packaging.[6] APHIS has amended its regulations several times since 2009. We urge the Subcommittee to call for APHIS to facilitate independent analysis of the efficacy of its current phytosanitary programs in order to understand whether the updated regulations have reduced the risk of additional introductions.
Again, pests introduced via this pathway proliferate and spread – often facilitated by movement of firewood, plants, and outdoor household goods. APHIS’ programs have suffered severe failures to prevent such spread, for example in the cases of the emerald ash borer and sudden oak death. We suggest that the Subcommittee inquire of APHIS what steps it will take to improve containment efforts regarding damaging plant pests, including through collaboration with its state partners.
We ask for small increases to the Pest Detection and Methods Development programs. The first enables prompt detection of newly introduced pests … which is critical to successful pest eradication or containment. The second empowers APHIS to improve essential detection and eradication tools.
The current emergency fund of is far below the level needed to respond when a new pest is discovered. We thank both the House and the Senate for clearly recognizing that these appropriations are inadequate by including in their bills language reiterating the Agriculture Secretary’s power to access funds from other Departmental programs (usually the Commodity Credit Corporation) to respond to emergencies.
[1] Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
[2] Aukema, J.E., B. Leung, K. Kovacs, C. Chivers, K. O. Britton, J. Englin, S.J. Frankel, R. G. Haight, T. P. Holmes, A. Liebhold, D.G. McCullough, B. Von Holle.. 2011. Economic Impacts of Non-Native Forest Insects in the Continental United States PLoS One September 2011 (Volume 6 Issue 9)
[3] Hudgins, E.J., F.H. Koch, M.J. Ambrose, and B. Leung. 2022. Hotspots of pest-induced US urban tree death, 2020–2050. Journal of Applied Ecology
[4] Jetter, K. A. Hollander, B.E. Nobua-Behrmann, N. Love, S. Lynch, E. Teach, N. Van Dorne, J. Kabashima, and J. Thorne. 2022. Bioeconomic modeling of invasive species management in urban forests: final report.
[5] Haack RA, Hardin JA, Caton BP and Petrice TR (2022) Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
[6] Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Congressional Committees with Jurisdiction … & how to submit testimony
FUNDING APHIS
House Committee on Appropriations, Subcommittee on Agriculture, Rural Development, Food and Drug Administration, and Related Agencies
Chairman: Andy Harris (R-MD)
Members: Robert Aderholt, David Valadao, John Moolenaar, Dan Newhouse, Julia Letlow, Ben Cline, Ashley Hinson, Scott Franklin
Democrats à Sanford Bishop, Jr., Chellie Pingree, Lauren Underwood, Marie Gluesenkamp Perez, Marcy Kaptur, Debbie Wasserman Schultz
deadline: May 1; email to ag.approp@mail.house.gov
instructions: 5 pages, double-spaced in Times New Roman, 12 Point Font; single-sided; PDF attachment to your email. At top of 1st page, clearly indicate your name, title, & institutional affiliation (if any); In 1st paragraph, clearly state agency, program, & amount of funding in the request
Senate Committee on Appropriations, Subcommittee on Agriculture, Rural Development, Food and Drug Administration, and Related Agencies
Chairman: John Hoeven (R-ND)
Members: Republicans à Mitch McConnell, Susan Collins, Jerry Morn, Cindy Hyde-Smith, Deb Fischer, Mike Rounds
Democrats à Jeanne Shaheen, Jeff Merkley, Tammy Baldwin, Martin Heinrich, Gary Peter, Kirsten Gillibrand, Jon Ossof
deadline: not clear; might be 22 May; email to agri@appro.senate.gov
instructions: 4 pages.. At top of 1st page, clearly indicate your name, title, & institutional affiliation; state agency, program, & amount of funding in the request
FUNDING USFS
House Committee on Appropriations, Subcommittee on Interior, Environment and Related Agencies
Chairman: Mike Simpson (R-WY)
Members: Republicans à Mark Amodei, Guy Reschenthaler, Michael Cloud, Ryan Zinke, Jake Ellzey, Celeste Maloy
Democrats à Chellie Pingree (D-ME), Betty McCollum, Josh Harder, James E. Clyburn
deadline: 22 April; email to IN.Approp@mail.house.gov
instructions: 4 pages, single-spaced in 12 Point Font; single-sided; prefer PDF but other formats OK. At top of 1st page, clearly indicate your name, title, & institutional affiliation (if any); In 1st paragraph, clearly state agency, program, & amount of funding in the request
Senate Committee on Appropriations, Subcommittee on Interior, Environment and Related Agencies
Chairman: Lisa Murkowski (R- AK)
Members: Republicans à Mitch McConnell, Shelly Moore Capito, John Hoeven, Deb Fischer, Mike Rounds
Democrats à Jeff Merkley, Chris van Hollen, Martin Heinrich, Tammy Baldwin, Kirsetn Gillibrand, Jon Ossof
deadline: unclear; possibly mid-June; email to int@appro.senate.gov
instructions: 4 pages, single-spaced in Microsoft Word or Word Perfect; do NOT send PDF. At top of 1st page, clearly indicate your name, title, & institutional affiliation (if any); In 1st paragraph, clearly state agency, program, & amount of funding in the request
In a press release on 31 March, 2026, the USDA announced major changes to the USFS structure. Agency headquarters will be moved to Salt Lake City. They point out that nearly 90% of USFS land is west of the Mississippi … but promise to sustain engagement in the Southeast (America’s “wood basket) by creating a regional office there. Furthermore, they will change the current regional organization to a state-based one; they plan to create 15 state directorships. State directors will serve as national leaders with primary oversight of forest supervisors, operational priorities, & relationships with states, tribes, & other partners. Each state office will include a small leadership support team responsible for functions such as legislative affairs, communications, & intergovernmental coordination.
There will still be some “operational service centers” in other cities; that for research will be in Fort Collins. The goal is to unify research priorities, accelerate the application of science to management decisions, & reduce administrative duplication. Information on which facilities will be retained or closed is available at this webpage. (I could not open this site.)
No specific information is provided re: forest health management program.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The US Department of Agriculture (USDA) and the North American Invasive Species Management Association (NAISMA) held the 34th annual forum on invasive species research at the end of February 2026. The agenda is available here. In this blog I summarize the presentations about invasive alien plants (IAS); a separate blog discusses findings on invasive plants. Formal proceedings will be available in some months.
The most important information from the meeting:
If NAISMA had not taken on the task of hosting the conference it would not have happened.
Government leaders allowed only 1 staffer per USDA Forest Service region to participate. Not allowed to come were people who had organized the whole meeting or individual sessions, and presenters discussing several topics, including preventing IAS plant spread, and progress on controlling cogongrass (major impediment to pine plantations, affecting harvests).
What do these decisions say about the genuineness of the USDA Secretary’s recent memorandum listing invasive species as one of four priority areas for the department’s research efforts?
The USFS International Program is one of the few sources of support for studying potential pests before they invade the US.
Early detection surveillance is undermined by reliance on deploying too few traps and in a too narrow, or the wrong, timeframe.
The Resistance Screening Center in Asheville, NC is no longer staffed, undermining breeding efforts in a region that reaches from Virginia to Texas.
A reminder to us all: Rebekah Wallace of the Center for Invasive Species and Ecosystem Health at the University of Georgia urged us all to provide citations for images used in informal materials – posters, presentations, outreach efforts, blogs, videos. Providing the citation increases our credibility and ensures that we avoid perpetuating mis-information!
an ash resistance breeding plot at the Holden Arboretum, Ohio
Summary of key research reports on tree-killing arthropods and pathogens
Jennifer Koch, researcher with the USFS, described the Trees in Peril program. TiP aims to increase the pace, scale and efficiency of resistance breeding programs for American beech; eastern hemlock; and green, white, and black ash. This includes integrating genomics with other approaches and strengthening partnerships. Partners are key to finding “lingering” trees, addressing some scientific questions, and possibly screening cuttings for resistance.
Koch first explained the value of resistance breeding for producing resistant stock for restoration and reducing habitat for pests. The goal is to develop resistance, which Koch defined as the ability of a tree to survive despite the pest. Full immunity is not required. TiP participants hope that by integrating breeding with other approaches, such as biocontrol, they can create a new ecological equilibrium in which the tree species will continue to play its ecological role. As Koch asserts, the public supports breeding more than some other approaches. Also, there is a record of success; she cites the USFS Dorena Genetic Resource Center, which has developed resistant seedlings for four five-needle pines and Port-Orford cedar.
The first step is to determine whether desired traits are inherited. Genomics and other tools can test cuttings while they are still young and small – a very important advance in efficiency. Still, once cuttings with the desired traits are identified, it often takes several rounds of breeding to raise resistance levels sufficiently high. Similar testing of immature clones later in the process also can accelerate creation of seed orchards.
Breeding programs also need to incorporate genetic diversity from across the species’ ranges. TiP partners are collecting genetic material from beech, hemlock, and ash trees across their extremely large ranges – much of eastern North America.
Finally, TiP is training additional people to contribute to these breeding efforts.
Progress on each taxon:
beech grafts in a breeding experiment at the Holden Arboretum
Beech – Breeders are dealing with two diseases. A decade ago they identified genetic markers associated with beech bark disease (BBD). Their efforts had led to orchards producing seedlings of which 50% are resistant. Then beech leaf disease (BLD) showed up! Early results of a pilot study suggest BLD symptom severity is under genetic control. Even better, some trees appear to be resistant to both diseases. Koch recommends that scientists first identify BBD-resistant trees, then test those trees for BLD resistance.
Ash – the emerald ash borer (EAB) is established in 40% of the range of ash species. (Note: I am not sure whether this statement includes Canada; I am fairly certain it does not include Mexico.) Nine of the 16 US species are vulnerable, five endangered – green, white, black, blue and pumpkin.
The process by which scientists determe that resistance traits are heritable and identifying promising genotypes is described in Mason et al. (2026). The effort to develop techniques to propagate rooted cuttings is described in Merkle et al. (2022).
Partners are helping to search for “lingering” ash. So far, 265 trees have been identified, and scion collected from 106 trees. Partners are also helping to plant cuttings for resistance testing.
The program has had to overcome several difficulties, including:
Black ash is dioecious, which complicates selection. Breeders are working on several approaches, but all are at early stages.
Many of the originally collected trees turned out to be unintended crosses of white and green ashes rather than pure species. This resulted in very low seed production.
Anticipating the introduction of ash dieback disease (caused by the fungus Hymenoscyphus fraxineus), TiP is collaborating with Europeans on searching for possible resistance to this threat as well.
Hemlock – the Hemlock woolly adelgid (HWA) causes mortality of 50 – 100% of overstory trees. TiP scientists are still trying to establish a test for heritability of HWA resistance. There are additional difficulties in propagating rooted cuttings. The University of Georgia, Holden Arboretum, and others are helping to resolve these issues.
Those who want to support this program by contributing funds, knowledge, facilities, or volunteer efforts should contact Dr. Rachel Kappler, Forest Health Collaborative Coordinator, Holden Forests & Garden.
One entity already actively helping the TiP program is the Ecological Research Institute through energizing citizen scientists. Radka Wildova described these efforts. The Monitoring and Managing Ash [MAMA] initiative has published detailed guidance on identifying “lingering” ash. For example, timing is crucial: searching too early points to trees that are not actually resistant. Searching too late means opportunities are missed (since “lingering” ash will die eventually because resistance is only partial) or a risk of confusing in-growth or regeneration for “lingering” trees.
The Institute could not create a similar action map for hemlocks because the adelgid has been present far longer. Recommends searching in sites where at least 80% of surrounding trees are dead or dying due to HWA or elongate hemlock scale. The program is also testing heritability of resistance among hemlocks on its own property, which was invaded 20-30 years ago.
[An unrelated initiative, the Hemlock Restoration Intiative, is pursuing protection and breeding efforts in the southern Appalachian mountains.]
Avalon Miller, Pennsylvania State University, discussed new techniques to detect American elm trees tolerant of this disease.
a healthy American elm in Fairfax County, Virginia; photo by F.T. Campbell
It is important to detect elm trees’ response to infection early in the infection process because the apparent mechanism of tolerance is some trees’ ability to limit growth and proliferation of the causal fungus Ophiostoma novo-ulmi in xylem vessels. Scientists sought to use spectral analysis to detect distal leaf stress as a signal of susceptible genotypes. The USFS has developed a small stem assay that is achieving 80% accuracy in identifying disease phenotype within two months of inoculation – before symptoms appear.
Future studies will focus on determining which metabolites vary in tolerant vs. susceptible trees, and whether that information suggests useful interventions. For example, it is thought that some trees respond too aggressively to the pathogen, thereby cutting off the flow of water and nutrients and killing themselves.
Meanwhile, continuing efforts to breed resistant elm are hampered by limited greenhouse space, the tree’s complex genetics, and vast geographic range, and great variation in trees’ responses.
Current USFS- and The Nature Conservancy-supported programs focus on the Northeast. I urge scientists in the Mid-Atlantic to engage; I have seen numerous healthy American elms in the Virginia and Maryland suburbs that could be included in a breeding program.
Courtney Johnson, North Carolina State University, described efforts to determine key aspects of the ALB invasion in South Carolina. First, the bad news: a second invaded site in the Charleston region was detected in 2025.
Because Charleston is much farther south than any other ALB infestation, questions have arisen about
its phenology (timing of development). Research has confirmed that the ALB in South Carolina has ~1 year development cycle, not multiple generations as some had feared. Beetle larvae stay in the phloem through the third instar. Adult flight season is from May – Sept; the peak is in July. Unlike earlier findings, adult beetles did not exhaust their natal tree before moving to a new tree to oviposit. (This is also true in the Massachusetts outbreak.)
Some of the beetles in South Carolina are larger. Outreach materials need to be amended to reflect this fact, e.g., much larger exit holes.
typical site of ALB infestation in Charleston South Carolina; arrows indicate infested red maple trees. Photo by David Coyle
Tree dissection and dendrology studies of the principal host, red maples, show that multi-stemmed trees and smaller branches are preferred. They also preferred vertical stems or bolts, although they did oviposit on horizontal bolts raised off the ground to mimic a tree branch. There was little oviposition on bolts on the ground. In practice this means managers can leave felled trees on the ground without prolonging the infestation. This is very helpful since swamps preclude using heavy equipment. picture
Chad Rigsby, Bartlett Tree Research Laboratory, described the results of testing the efficacy of several nematocides. A foliar spray, Bayer’s Broadform, has received emergency approval from many states. It suppresses nematode (Litylenchus crenatae mccannii ([LCM]) numbers when applied at very low rates. Trees can be treated as long as (green) leaves are present. Rigsby recommended not spraying until a tree displays symptoms.
Since foliar sprays cannot be applied in forests, near water, or on huge trees, scientists also sought a systemic injectable fungicide. Thiabendazole [TBZ] (commercial formula Arborjet 20-S) is available. Rigsby said applicators can avoid splitting of the bark by following protocols developed by the International Society of Arboriculture. Managers should inject a tree several times in the first year to get the disease under control; then they can apply less frequently.
injection of thiobenzadole into beech; photo by Matthew Borden of The Bartlett Tree Research Laboratories
Don Grosman of Arborjet believes mortality is the result of a disease complex, not just LCM. Any of three treatments containing phosphite greatly reduces nematode numbers and canopy symptoms. Low volumes of diluted product can be injected in a few minutes. However, Thiabendazole hypophosphite requires a high volume macro trunk injection. This is expensive and takes time
Testing shows potassium phosphite PHOSPHO-jet produced dramatic improvement in 1 year. There are early indications that one treatment might be effective for two years. Arborjet will test this finding again this year. The company is also testing another chemical – the name of which cannot yet be revealed.
Andrew Miles, Ohio State, described beech response to polyphosphate (PP30). This chemical is a biostimulant, not a treatment. It is used as a disease control agent in several crops, including woody species. Field observations indicate it does reduce disease severity. Scientists are trying to understand the mode of action. Experiments are under way in Cleveland MetroParks, where BLD was first detected. Miles called for experiments within buds as well as leaves, since LCM damages tissue while in the bud.
Scott Schlarbaum, University of Tennessee, collects butternuts; photo by F.T. Campbell
Anna Conrad, USFS, described ongoing efforts targetting this disease, which is present throughout the tree’s large range. A major challenge is distinguishing pure butternut from hybrids with Japanese walnut. Scientists have screened ~300 families from 22 states for possible resistance. At three sites in Indiana, the vast majority of highly resistant families are hybrids. Still, resistance was detected in up to 2.5% of pure butternuts; this level is sufficient to be enhanced through breeding. The program would benefit from genotyping across butternut’s range to identify lingering trees and confirm resistance.
Nicholas Dietschler, Cornell University, studies the relationship between western hemlocks and HWA in their shared native ranges in the Pacific Northwest. At all sites, lower numbers of HWA (of both PWN and Japanese lineages) survived on Western hemlock – in the absence of predators. Why? Dietschler believes western hemlock has better chemical defenses. For example, hemlocks exude pitch in response to adelgid herbivory. In eastern hemlocks, this induced resin might suppress the tree’s defenses. In addition, HWA also prompts greater suppression of phenolics in eastern hemlock. Dietschler concludes that bottom-up, tree-based defenses are a factor in the invasion and should be studied — while continuing efforts to find an effective combination of biocontrol agents.
Anne C.J. Peter, of Virginia Polytechnic Institute and State University, is comparing HWA chemical interaction with the most recent biocontrol agent, the silver fly Leucotaraxis argenticollis. (Scientists hope L. argenticollis will feed on summer populations of HWA; other biocontrol agents don’t suppress HWA at this stage.) The L. argenticollis population in the PNW feeds on HWA; however, its eastern North American relative L. rubidus feeds on pine adelgids, not the introduced HWA. It has been challenging to establish the PNW population in the East. One possibility is that the invasive HWA, which is from Japan, contain toxins that deter predators & parasitoids. Therefore, Peters is studying how both the western and eastern populations of Leucotaraxis deal with anthraquinones – compounds found in many plants and some insects, but not adelgids native to the eastern US.
Jian Duan, of the Agriculture Research Service Beneficial Insect Lab, summarized results of 15 years of biological control efforts. Over this period, four biocontrol agents have been introduced. I applaud APHIS’ rapid inclusion of this pest management approach; an egg parasitoid and two larval parasitoids were introduced before 2010, less than 8 years after the invasion was detected. Unfortunately, these agents proved less effective in northern parts of the EAB’s distribution. A fourth larval parasitoid was released in 2015. One or more of these biocontrol agents have been released in 479 counties in 34 U.S. states and three Canadian provinces.
To what degree have the wasps reduced EAB populations? Are those reductions resulting in regeneration?
Duan reported that at sites in Michigan, all four agents have spread rapidly. EAB populations crashed and recovered several times but overall numbers are lower. Ash saplings increased greatly after 2015; seedlings also increased. He concluded that the program has been successful but not spectacularly so.
Hannah Broadley, APHIS, described developments beginning with initial searches for possible agents in China in 2015 — just one year after the lanternfly was detected in Pennsylvania. The search has focused on agents that feed on SLF eggs and nymphs. Attention has narrowed to Dryinus sinicus. This wasp both preys on and parasitizes SLF nymphs – depending on the nymphal stage. Labs are developing a third colony and conducting host specificity testing. Scientist have begun drafting a petition for release; the review process will probably take more than one year. At the same time, scientists continue exploring other possible biocontrol agents – e.g., in Vietnam. The blizzard prevented this speaker from appearing.
Xingeng Wang, of the ARS Beneficial Insect Lab, described how Dryinus sinicus attacks SLF – with a graphic video! D. sinicus attacks on third instar are often unsuccessful. When it encounters a second instar nymph, however, D. sinicus switches from predation to parasitism: it lay an egg which then develops inside the SLF nymph. This parasitism kill seven times more nymphs than predation on older nymphs.
Individual D. sinicus wasps can live up to 60 days, lay an average of 175 eggs and parasitize ~137 nymphs! Since D. sinicus is most effective against just one instar, releases will need to be carefully timed.
Alex Wu, APHIS, discussed efforts to prevent establishment of four flighted spongy moth (Lymantria) species. APHIS seeks to improve the efficiency of trap analysis because states are submitting triple the number of trap contents of past years. The goal is to improve real-time qPCR efforts to distinguish the European species established in the East from the Asian flighted species, and to distinguish the several subspecies of latter taxon. Current qPCR results point to the wrong species ~ 5% of time. There are complexities: moths from Central Asia might be hybrids. Also Lymantria dispar japonica might be found in far southeastern corner of Korea – which is separated from Japan by a narrow strait.
Early Detection of Wood-Associated Beetles
Jiri Hulcr, University of Florida, discussed strengths and weaknesses of artificial intelligence (AI) in species identification of bark beetles. As he noted, differentiating a specific bark beetle species from among the more than 6,000 look-alike taxa is time-consuming. A properly trained AI program can help. Furthermore, no one can keep up with publications – in 2015 there were 432 discussing just bark beetles! AI can help researchers discover papers that they otherwise would miss and empower non-English speakers to search the literature.
Hulcr has created a website that now has 63,000 images of ambrosia and bark beetles to assist identification. This work has been funded by the USFS International Program – one of the few sources of support for studying potential pests before they invade. The website will be open source once it has been copyrighted to prevent “scraping” by bots. Hulcr invited participants to send more images to continue training the algorithm on more species.
In the discussion, Alain Roques noted that scientists in Europe and probably China are developing similar AI-assisted identification tools. He urged international coordination. Hulcr replied that scientists do coordinate – as long as funding is available. Jennifer Koch noted that historic collections have many taxonomic inaccuracies. She urged people to rely on genetics when trying to identify a species.
Hulcr says AI is much faster than people in completing some tasks. But managing bioinvasions continues to require trained people (taxonomists) to collect, detect and classify new species; and execute quality control. AI cannot do science, which Hulcr defined as generating new knowledge through observation, turning that information into data, and testing hypotheses, making assumptions based on that.
Hulcr says AI also cannot predict what the next damaging ambrosia beetle to enter the U.S. will be. He offered his predictions:
Euwallaceae destructans – from Indonesia – attacks live trees
Aggressive Platypdinae from Asia and South America (especially threatening to plantations where trees are stressed)
Cryphaus lipingensis (attacks pine seedlings)
Scolytus amygdali from the Meditteranean region – introduces pathogens during maturation feeding on living hosts; feeds on almonds and prunes – Rosaceae
Dryocoetes himalayensis – Asia and Europe; kills walnuts
Port of Marseille; via Wikimedia
Alain Roques, of Zoologie Forestiere in France, reported results of a beetle trapping study in France.
Since the European Union allows entry of species not listed as quarantine pests, it is vitally important to improve detection and analysis of the large percentage of detections that are “unknown” or “emerging”. Nearly 8,000 beetles have been trapped over five years; they belong to nearly 400 species, 35 non-native.
One approach is to develop more generic traps and lures. The EU is now using a blend combining 10 pheremones to trap Cerambycidae. Scientists are incorporating additional pheromones to the blend and to extend attractiveness longer than the current 10 days. There is still no generic lure for Buperstids.
Some species arrive regularly – is each detection a reintroduction? Or are these species established?
Roques asks whether we are trapping at the right sites. Half of Cerambycids are trapped only inside ports (of various types). Scolytids were trapped outside ports, at other “high-risk” locations– e.g., sawmills and recycling centers. In other words, they disperse more broadly. Roques wants to add the road network and to extend the survey to the entire European Union.
Davide Nardi, of the University of Padua, Italy, discussed results of his trapping program, which seeks to guide placement of traps. See Nardi et al. (2026) [full citation at end of this blog]. Important conclusions are:
Surveillance programs are probably under-sampling species. Halving the sampling effort (from 16 to 8 traps) resulted in failure to detect ~20% of the species at the site. Cutting the sampling effort to four traps resulted in missing ~ 40% of present species. This decline in catches is particularly severe in urban landscapes – the very places where insects are most likely to be introduced. Even when they deployed 16 traps per site almost 30% of total species richness was not detected, on average.
Urban landscapes might offer a higher diversity of potential tree hosts. They also have more barriers to insects’ spread, e.g., buildings. This means urban areas require a greater sampling effort.
Traps should be set near available forest patches or urban parks.
I was intrigued by Nardi’s suggestion that scientists use the data on native beetles included in the trap catches to alert countries receiving exports from these ports to which species might be transported to their shores.
Manoj Pandey, of Ohio State University, explored how environmental context shapes abundance and diversity of Scolytines caught in surveillance traps. His goal is to improve the efficacy of the USFS’ two- decade-old Early Detection Rapid Response trapping program, which targets bark and ambrosia beetles at high-risk sites. These include transit sites, destination sites, and wood waste treatment sites. Pandey analyzed program catches from 2010 through 2019.
He found that among native species bark beetles dominated catches; ambrosia beetles dominated non-native captures. Climate [minimum/maximum temperature and precipitation] was the most important factor determining which species were caught. Overall, both Scolytines and ambrosia beetles are governed more by ecological requirements than by human population levels. Among Scolytines, native species (which are adapted to stressed trees) are affected by precipitation; non-native species are favored by warmer temperatures. Ambrosia beetles – both native and non-native – are more affected by precipitation levels than bark beetles, probably because of the formers’ symbiotic relationship with fungi. Ambrosia beetles are also more likely to be generalists and to be attracted by deciduous forests.
The other influential criterion was landscape – whether forests are evergreen, deciduous, or diverse. Deciduous forests attract both types of beetles, but the influence is stronger for non-natives. Conifer (evergreen) forests attracted native species. Higher human population density was associated with higher trap catches. Propagule pressure – measured via human population density and per capita income – was less important, perhaps because the traps are always placed near population centers.
Xyleborus monographus; photo by U. Schmidt
I am concerned because this trapping program did not detect the Mediterranean oak borer (Xyleborus monographus) before it was detected in California and Oregon. The project also did not find the greater shot hole borer Euwallaceae interjectus on the West Coast before it was detected in Santa Cruz, California. This ambrosia beetle has been established in the Southeast for years (M. Pandey, pers. comm. 12 March 2026).
Other Pests and Pathogens
Thomas G. Paul, at Ohio State, explored whether understanding the temperature regime during transit can provide early warning of which wood-associated pests might arrive. He obtained ocean surface temperature data along shipping routes from China to the U.S. West Coast and across the Atlantic. He then related those temperatures to degree-days needed for development by Xylosandrus germanus (from Asia) and Ips typographus (from Europe). At present there is still lots of uncertainty, including how to factor in the insect’s stage at the time of departure, the relationship between ocean air temperature and temperature inside a container, and possible effects of a container left to sit for several days in the port of import.
Eliana Torres Bedoya, also of Ohio State, provided an update on spore trapping for improved detection of pathogens across large landscapes. In 2024 the project developed standardized protocols for surveillance. To learn what is going on in the region, one should sample many sites across the area of interest. To find a particular pathogen, officials also need to know which season to sample. Torres Bedoya notes that few states sample in the autumn, which probably results in biased results.
In 2025, the program was expanded to 10 states. Species searched for are chosen by participating states. They include the causal agents of oak wilt, thousand cankers disease, laurel wilt, annosum root disease, and the beech leaf disease nematode (Litylenchus crenatae mccannii). Participants – including state phytosanitary officials — are now asking how to respond to a detection. For example, DNA from Bretziella fagacearum, the cause of oak wilt, was detected in several states where no disease has yet been identified (New Hampshire, Massachusetts, West Virginia, and Ohio). DNA of Geosmithia morbida, the causal agent of thousand cankers disease of walnut, was detected in New Hampshire, Massachusetts, and Maryland. What should managers do in response to these findings?
Torres Bedoya explained that her team is now working to make the spore-trapping process more user-friendly. I noted that my poster previous blog discussed using these techniques at the interface of forests and agricultural land uses.
During other discussions, I learned that Jason Smith of the University of Mount Union is trapping for DNA from LCM in order to track the spread of BLD.
Brown spot needle blight
Several speakers addressed this disease, which is of increasing concern to pine timber interests in the American South and around the world. New Zealand is exploring resistance breeding of Pinus radiata in advance of introduction
The disease has long been known in long-needle pine – at the “grass” stage (early seedlings). In recent years needle blight has begun damaging loblolly and other pine species – in both plantations and natural forests. Jason Smith, from the University of Mount Union, was asked for help by the industry in 2016. He found that one factor is increasing reliance on herbicides instead of fire to control ground-level vegetation. The large doses of inoculum remains in the litter, rather than being killed by periodic fire – as in the past. Smith thinks it is also possible that the pines suffer subtle damage from herbicides. Other possible factors are the widespread planting of genetically identical monocultures and climate change.
Colton Meinecke at the University of Georgia reported that Lecanostica acidola has been confirmed as the disease agent at these sites by Koch’s postulates. Scientist at the University of Georgia, University of Mississippi, and other entities are collaborating on development of a predictive model. Work includes sampling needles from both the litter and canopy, tracking tree condition, destructive sampling of dead trees, and spore trapping.
In the discussion, Smith warned that dying pines are not being detected by aerial forest health surveys because they are conducted too late in the season. This is because the surveys focus on one specific pest, the southern pine beetle. He called for a more comprehensive survey program.
Meinecke reported that the disease is more severe in western parts of the Gulf Coast regions. It is also causing problems in Christmas tree plantations, especially Scots pine.
He has found evidence of some genetic resistance. He is trying to develop a rapid test of a tree’s vulnerability using spectral wave length. Meinecke is also experimenting with stand management approaches. He praised the close cooperation with experts from around globe and New Zealand’s pro-active preparation for combatting the disease before it arrives.
Kier Klepzig, of the Jones Center at Ichauway in Georgia, described establishment of a Pine Pandemic Preparedness Plan, stimulated by awareness that a non-native pest might be introduced that attacks loblolly pine (Pinus taeda) – the foundation of the southeastern “woodbasket”. [Of course, Sirex noctiliois already established in the eastern United States. Although it is a severe pest of loblolly growing in plantations in the Southern Hemisphere, industry and federal and state agencies have dismissed concerns in North America.] The Pine Pandemic Preparedness Plan has four components: communication, detection and diagnosis, delimitation and assessment, response.
As concern about brown spot needle blight grew, the Southern Group of State Foresters ask the “P4” team to engage. Klepzig and Kamal Gandhi pulled together a working group which has the goal of developing guidance for managing the disease within two to five years. The task force is developing a website for data-sharing. The task force is also studying genetics of the host and pathogen, fungicides, the role of fire, resistance screening, and spore trapping. Industrial concerns about coordinating with competitors cause challenges.
Ashley Schulz, of Mississippi State University, has reviewed experience with biocontrol for clues on species’ traits important for facilitating invasion. She analyzed information on 394 insects introduced to North America for biocontrol of invasive plant species (see other blog) and 87 agents targeting 325 insect pests. For each species, data was recorded on whether it established, level of impact, the insect’s feeding guild, climate matching, host specialization, and evolutionary history. For the 87 entomophagous insects, she also recorded host feeding guild and host specialization.
Schulz found that entomophagous insects introduced as biocontrol agents were more likely to establish if they are a specialist. Higher impact was also associated with specialization. Parasitoids had higher impacts than predators. What does this indicate re: invasive species? Schulz said that insects which can hide or defend themselves, i.e., specialists, are likely to be more successful invaders.
Schulz recommends more analysis of what can be learned from experience with biocontrol agents. However, such studies are challenged by poor records, lack of empirical evidence and quantitative data, the lower number of biocontrol agents introduced recently, and funding shortages that preclude post-release monitoring.
Schulz also mentioned that she worries that a proposal to drop the word “harm” from definition of invasiveness could result in biocontrol agents being lumped with invasive species. This would further hamper implementation of biocontrol. She considered this loss to have particularly bad affects at a time when there are growing restrictions on pesticide use.
SOURCES
Mason, M.E., Carey, D.W., Romero-Severson, J. et al. Select genotypes of white and green ash show heritable, elevated resistance to emerald ash borer. New Forests57, 12 (2026). https://doi.org/10.1007/s11056-025-10158-x
Merkle, S.A., J.L. Koch, A.R. Tull, J.E. Dassow, D.W. Carey, B.F. Barnes, M.W.M. Richins, P.M. Montello, K.R. Eidle, L.T. House, D.A. Herms, K.J.K. Gandhi. 2022. Application of somatic embryogenesis for development of emerald ash borer-resistant white ash and green ash varietals. New Forests https://doi.org/10.1007/s11056-022-09903-3
Nardi, D., D. Rassati, A. Battisti, M. Branco, C. Courtin, M. Faccoli, N. Feddern, et al. 2026. “Integrating Landscape Ecology into Generic Surveillance Plans for Bark- and Wood-Boring Beetles.” Ecological Applications 36(2): e70194. https://doi.org/10.1002/eap.70194
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm