Posted by Scott J. Cameron, former Acting Assistant Secretary for Policy Management and Budget , US Department of the Interior
locations in Hawaii Volcanoes National Park where ohia trees were infected by rapid ohia death pathogen in 2017
In 2019 Congress passed the John D. Dingell, Jr. Conservation, Management, and Recreation Act. Among other things, it directed the Department of the Interior to “develop a strategic plan that will achieve, to the maximum extent practicable, a substantive annual net reduction of invasive species population or infested acreage on land or water managed by the Secretary.” This provision triggered a year-long process of public involvement and inter-bureau coordination. The result was a plan published in January 2021. It represents the first attempt by Interior at a Department-wide multi-taxa, multi-year approach to invasive species. It has the potential to spur integration of invasive species work across the Department’s many bureaus and to focus each bureau’s efforts on a set of common goals, strategies, and performance metrics.
While Congress’ language is open-ended, any planning exercise is constrained by the most recent President’s budget and existing law. It is up to those of us who are not, or at least no longer, executive branch employees to advocate for plans unbound by those constraints.
In that spirit, I offer eight recommendations to improve invasive species management. Four are within existing authorities; four more are outside the current budgetary and statutory framework. Many more ideas are without a doubt worth pursuing.
Opuntia (prickly pear) cactus – common plant in western National parks and on Bureau of Land Management lands; under threat by cactus moth
First, within existing funding and legal authorities, the Department has unfinished business that it can act on now.
Secure approval of the package of categorical exclusions under the National Environmental Policy Act (NEPA) — now awaiting approval by the President’s Council on Environmental Quality (CEQ). Over many years agencies have documented how invasive species control improves, rather than harms, the environment. Absent an applicable NEPA categorical exclusion approved by CEQ, though, each such action needs to run the time- and money-consuming gauntlet of NEPA compliance. In the meantime, the invasive plants germinate, the invasive animals reproduce, and what might have been a localized and inexpensive problem has expanded geographically and in terms of cost and complexity. In the Fall of 2020 Interior submitted hundreds of pages of documentation on numerous practices proven to control invasive species without harming the environment. Due to time and staffing constraints at CEQ, these categorical exclusions still await action. Interior and CEQ should take prompt steps to finish them.
Allocate to the US Geological Survey at least $10 million of the roughly $90 million remaining available to the Secretary in CARES Act appropriations for research on invasive zoonotic diseases. These are diseases like COVID-19 and West Nile virus that can move from one species to another. Many zoonotic diseases fit the definition of invasive species, since they are not native to the United States and endanger human health. In the case of COVID-19 funding is available to the Secretary of the Interior, without need for further Congressional action, from funds appropriated by the CARES Act. The availability of these funds will expire at the end of September 2021, so Interior should provide this research funding to USGS as soon as possible.
Join the existing Memorandum of Understanding between the Western Governors Association (WGA) and the US Department of Agriculture’s Forest Service. These groups have established “a framework to allow the U.S. Forest Service (USFS) and WGA to work collaboratively to accomplish mutual goals, further common interests, and effectively respond to the increasing suite of challenges facing western landscapes.” This provides a forum to improve coordination between the States and the federal government on the management of invasive species, and Interior needs to be part of that team.
Interior should work with USDA to accelerate and intensify efforts to systematically improve coordination between the interagency Wildland Fire Leadership Council and the interagency National Invasive Species Council. Both Councils have member agencies that practice vegetation management using similar tools and techniques, although for different purposes. The two Councils should identify a select number of initiatives in FY21 where their efforts would benefit both wildland fire management and invasive plant management .
swamp bay trees in Everglades National Park killed by laurel wilt; photo by Tony Pernas
Four steps to implement the strategic plan outside the scope of current law and the President’s Fiscal Year 2021 budget. I offer the following:
Improve implementation of the Lacey Act program to list injurious species. There are both legislative and administrative elements to this proposal.
In a federal District Court decision on May 19, 2015, on a lawsuit filed by the Association of Reptile Keepers, the Court undid the longstanding Fish and Wildlife Service policy that the Lacey Act allowed FWS to ban interstate transport of injurious species. On April 7, 2017, the D.C. Circuit Court of Appeals upheld the District Court’s view. These rulings mean that FWS authority only applies to international commerce. Unfortunately, the court’s interpretation of the law and legislative history are reasonable, so Congress needs to amend the law to make it clear that FWS is explicitly authorized to regulate interstate commerce in injurious species. The Department of the Interior should work with the Department of Justice and the Office of Management and Budget to develop the necessary bill language and submit it to Congress.
At the same time, the FWS injurious species listing process is notoriously slow, even causing Congress to occasionally list species legislatively. The fact that these legislative initiatives have sometimes been promoted by Members of Congress who normally are opposed to more federal regulation signals just how awkward the current FWS process is. Thus there might well be strong bipartisan support to amend the Lacey Act on the interstate commerce matter. Acting on its own authority, FWS should procure an independent third party review of the injurious species listing process and ask the contractor to make suggestions for “business process reengineering” to improve and streamline current practices, along with evaluating whether higher funding or new technology is needed.
The Bureau of Reclamation, Department of the Interior, and the Office of Management and Budget should develop legislative language to submit to Congress for the 2022 Water Resources Development Act, that explicitly authorizes an aquatic nuisance species program in the Bureau of Reclamation. It could parallel the relatively new authority enjoyed by the Army Corps of Engineers and mandate increased coordination between the two water agencies.
Any climate change legislation pursued by Congress and the Administration should include provisions for addressing invasive species. Climate change will make some North American habitats more suitable for foreign organisms, as cold-intolerant species might be able to survive in more northern latitudes in the U.S. than previously was the case. Second, the disruptive effect of climate change on North American ecosystem structure and trophic relationships at any latitude will make those ecosystems more vulnerable to invasion. Finally, the spread of invasive species may in and of itself exacerbate climate change, such as through the increased carbon dioxide emissions from rangeland wildfires aggravated by the dominance of invasive cheatgrass.
The Senate Committee on Environment and Public Works and the House Natural Resources Committee should each hold oversight hearings on how best to help state and local governments detect and respond to new invasive species that are not within the statutory purview of USDA/APHIS. This is a complex topic, important for both ecological and financial reasons. Over the years several attempts to address it have failed. A thoughtful review by Congress and the Administration, which perhaps would lead to passage of new statutory authority and funding, is worth exploring.
The Author:
Scott Cameron recently left the federal government, where he had served as Acting Assistant Secretary for Policy, Management and Budget at the U.S. Department of the Interior. In that capacity he oversaw Department-wide budget, invasive species, natural resource damage assessment and restoration, environmental compliance and numerous administrative functions of this $14 billion agency with 65,000 employees. Previously, Scott held other civil service job (e.g., at the White House Office of Management and Budget) & political appointments in the federal government, on the staff of the Governor of California, and on Congressional staffs. While not in government, Scott led formation of the Reduce Risk from Invasive Species Coalition.
CISP welcomes comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
Further information on the plant pests mentioned in the photo captions can be obtained at www.dontmovefirewood.org; click the “invasive species” button.
Japanese stiltgrass in Shenandoah National Park; photo by Jake Hughes, NPS
The recent appearance of a study by National Park Service (NPS) scientists quantifying the threat posed by invasive plants prompted me to seek updates on this issue. In May 2018, I blogged about NPS’ Invasive Plant Program Strategic Plan, which was issued in 2016. At the time, I got the impression that the program was struggling to gain support from NPS leadership in Washington, as well as leaders of individual parks. Has the situation improved since then?
The recent study, by Kathryn Miller and colleagues, focuses on National parks in the East, from Virginia to Maine. (I provide a full reference at the end of this blog.) I look forward to a planned follow-up article that will try to clarify drivers of invasion.
The Miller study appears to have been undertaken to partially fulfill one of the goals of the NPS’ 2016 Invasive Plant Program Strategic Plan. One of the actions under Goal 1 in the Plan was to quantify the invasive plant threat, the effort needed to manage it, and then to communicate the gap between needs and available resources. Miller et al. have quantified the threat to National parks in one region (not the entire country). However, they have communicated the gap between effort and need in only the most general way.
Protecting the forests in eastern National parks is valuable from many perspectives. The forests in these parks are older and have higher stand-level tree diversity than surrounding unprotected forests. Protecting their long-term condition also corresponds to the National Park Service’s (NPS) stated mission, as established in its Organic Act.
The study relied on National Park Service Inventory and Monitoring Program data from 1,479 permanent forest plots covering 39 eastern NP units located from Virginia to Maine. The data were collected over 12 years, in three survey cycles (2007–2010, 2011–2014, and 2015–2018).
The authors note that in forest ecosystems, invasive plants can reduce native diversity, alter forest structure, suppress tree regeneration, alter nutrient cycling, and modify disturbance regimes. Some – the shrubs and vines – also threaten human health by promoting increases in tick populations and associated Lyme disease risks.
Unsurprisingly, the data showed invasive plants to be widespread, and increasing. In 80% of the park units, there was a significant increase in at least one trend measuring abundance. Any decrease in plant invaders tended to be in herbaceous or graminoid species; that decrease was often counterbalanced by an approximately equivalent increase in invasive shrubs or vines. In 35 of the park units, more than half of the plots had at least one invasive plant species when the 2015-2018 survey began. In 10 parks (a quarter of those surveyed), every plot had at least one.
The most widespread species is Japanese stiltgrass (Microstegium vimineum). It is present in more than 75% of all park units and 30% of all 1,400 plots. This dominance is true despite the fact that stiltgrass does not extend north of Roosevelt-Vanderbilt National Historic Site in New York state (41o N latitude). Stiltgrass is also the most aggressive invader.
The second most widespread invaders are a group of shrubs and vines, including most notably Japanese barberry (Berberis thunbergii), Japanese honeysuckle (Lonicera japonica), multiflora rose (Rosa multiflora), and wineberry (Rubus phoenicolasius). I note – although the articles does not – that several of these have been deliberately planted – either to “enhance” ecosystems (multiflora rose) or as ornamentals (barberry). [Lehan et al. 2013 (full reference at end of blog) found that 95% of 125 shrub species introduced to the U.S. were introduced deliberately.] I know of no examples of deliberate planting of Japanese stiltgrass.
The most frequently detected non-grass herbaceous species is garlic mustard (Alliaria petiolata), which was detected in 20% of plots. Garlic mustard is found throughout the study area (Virginia to Maine). Tree-of-heaven (Ailanthus altissima) is the most common invasive tree. It is found in only 9% of plots and does not grow north of Roosevelt-Vanderbilt NHS. Again, both were intentionally introduced – and not noted as such in the article.
Because they could not identify the source populations for each plot, the study could not directly measure rates of establishment and expansion. The data did allow tracking rough trends in each park.
Parks with the highest abundance tended to be near densely populated areas. However, this pattern was not universal. For example, Prince William Forest Park in the outer Virginia suburbs of the District of Columbia metro area was one of two of the least invaded park units.
Prince William Forest Park
(note the prevalence of beech – I fear for the arrival of beech leaf disease!)
Total invasives increased significantly in 21 of the 39 parks for at least one metric. In 10 parks (a quarter of all parks), total invasives increased significantly in two of the three metrics. Antietam National Battlefield experienced the steepest increases.
The authors note that invasive plants continue to establish and expand, even in already heavily invaded forests. Thus they found little evidence of saturation. This finding conflicts with invasion theory. They also found antagonistic interactions between invasive species to be common.
The authors said managers should prioritize efforts to control Japanese stiltgrass and the shrubs and vinesdue to their widespread occurrence, rapid expansion, ability to suppress tree regeneration and understory diversity, and – in the case of the shrubs and vines, link to ticks.
The authors noted the need to better understand the drivers and impacts of invasive plants in eastern forests. They mentioned the overabundance of white-tailed deer (Odocoileus virginianus), latitude, climate change, fragmentation and urbanization. I have urged them to include analysis of deliberate planting of various species on lands within the park units or nearby. They have said that they will do so in the planned follow-up article.
The authors propose that deferred management of natural resources receive attention and sustained commitment on par with the attention to deferred maintenance of the park units’ infrastructure. The maintenance backlog has been addressed through recent legislation: the Great America Outdoors Act which provided $6.5 billion over five years to address deferred maintenance projects in all 419 national park units. I believe that these projects will not address invasive species, which are managed under the “Natural Resources” budget account, not “Maintenance”. Some “Maintenance” projects probably will include control of invasive plants. A former Interior Department official has suggested that resource management activities might be funded under another section of the legislation, which provides $900 million under the Land and Water Conservation program. I hope this is true.
National Park Service’ Overall Invasive Plant Program Now
In response to the mandate in the John D. Dingell, Jr. Conservation, Management, and Recreation Act (Public Law 116-9), the Department of Interior has issued a department-wide invasive species strategy. Will issuance of the new strategy provide impetus to the NPS to seek funding to implement its 2016 invasive plant strategic plan? Will Congress provide funds for this purpose?
Finding out the current status of National Park Service took a little effort. The NPS’ website has “popular” information about the efforts of individual regional Invasive Plant Management Teams (see links at the end of this blog). These posts provide only the briefest overview of program achievements and do not compare those accomplishments to the goals in the 2016 plan.
However, Terri Hogan, Invasive Plant Program Manager in the Biological Resources Division, provided following information:
NPS leadership now supports the agency’s invasive plant effort. The national Invasive Plant Program (IPP) contributes to the annual Natural Resource Stewardship and Science Directorate (NRSS)’ Biological Resource Division (BRD) work plan. All is guided by the “Four Pillars to Guide Natural Resource Activities and Investments”, adopted in 2016.
It is not clear that invasive species have the highest priority under this regime. The four “pillars” are
Holding the line – includes conserving biodiversity by removing invasive plants
Managing amid Continuous Change – includes conducting risk assessment and taking other actions to contain future exotic species
Leveraging for Conservation at Scale
Enhancing Stewardship and Science Access and Engagement
Cooperation with owners of neighboring private lands has been enhanced by engagement of the Western Governors Association and state and local political leaders. Many parks participate in Partnerships for Invasive Species Management (PRISMs), CISMS, and Cooperative Weed Management areas (CWMAs). This collaboration has been strengthened by adoption of the John D. Dingell, Jr. Act (see above).
In practice, the focus appears to still be on the Invasive Plant Management Teams (IPMTs). There are now 17 teams. Fifteen are funded through the national office. One is funded by an individual park; one funded through a regional office. Annual reports have been published for FYs 2017 and 2018. The FY19 report has been held up but should be posted soon.
The reports provide brief description of the overall program and vignettes of particular activities. There are more detailed – but still anecdotal – reports for each of the teams. It is difficult to determine whether there has been overall progress. For example, the reported total infested acreage increased from 133,658 acres in FY17 to 301,195 acres in FY18. This presumably reflects more intense monitoring as well as a probable increase in real infections. (The infested acreage figures do not address intensity of invasion on these acres.) The teams cumulatively treated 8,937 acres in FY 2017; 8,331 acres in FY 2018. They carried out inventory and monitoring projects on 169,057 acres in FY17, 210,000 acres in FY18.
Since the Miller article concerns the region from Virginia to Maine, I checked the FY17 and FY 18 reports from the Invasive Plant Management Teams from the Mid-Atlantic, National Capital Area, and Northeastern regions. The Mid-Atlantic team emphasized work on wavyleaf basketgrass and meadows in parks established to protect cultural or historic sites. The National Capital Area team emphasized partnerships and success eradicating Giant Hogweed (Heracleum mantegazzianum) on private land neighboring Rock Creek Park. The Northeastern Team serves 25 parks; the report emphasized leveraging resources and testing efficacy of pre-emergent herbicide for control of Japanese stiltgrass. With this exception, the teams do not appear to be prioritizing the species recommended for action by the Miller study.
Brief, “popular” articles about the NPS’ invasive plant effort are available here
Lehan, N.E., J.R. Murphy, L.P. Thornburn, and B.A. Bradley. 2013. Accidental Introductions are an Important Source of Invasive Plants in the Continental United States. American Journal of Botany 100(7): 1287–1293. 2013.
Miller, K. M., B. J. McGill, A. S. Weed, C. E. Seirup, J. A. Comiskey, E. R. Matthews, S. Perles, & J. Paul Schmit. 2020. Long-term trends indicate that invasive plants are pervasive and increasing in eastern national parks. Ecology. 00(00):e02239. 10.1002/eap.2239
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
EAB-kiled ash tree in Shenandoah National Park in 2016 photo by F.T. Campbell
As we have known for years, forests of the eastern United States are under severe pressure from non-native forest insects and diseases. Several recent studies have put this fact into perspective.
Fei et al. (2019) found that the 15 most damaging introduced species threaten 41.1% of the total live forest biomass in the 48 conterminous states. Nine of the 15 species included in this calculation are pests of the eastern forest. Indeed, the greatest increase in biomass loss, as measured by USDA Forest Service Forest Inventory and Analysis (FIA) plot data occurred here. Compensatory growth in unaffected trees and the recruitment of new regeneration occurs only later – as much as two or more decades after the pest invasions began. Fei et al. (2019) expect these losses will be exacerbated in the future due in part to the likelihood that additional pests will be introduced.
Randall Morin found that non-native pests had caused approximately 5% increase in total mortality, by tree volume, nation-wide.
Most widespread pest threats in the East
Scientists have used several methods of measuring introduced pests’ impacts. One measure is the number of counties where the pest is present. A second measure is the proportion of the volume of the host that has been affected. Both metrics are used by Morin. A third method, used by the CAPTURE Project (Potter et al. 2019a), is the number of hosts affected by the pest.
Morin and colleagues found that the European gypsy moth has invaded 630 counties – or 29% of the volume of its principal host, oaks. (In both cases, the gypsy moth trailed white pine blister rust in extent of infestation. The latter is nationwide but having its greatest impacts in the West). The CAPTURE Project found that the gypsy moth affected the largest number of hosts – 65.
Using the “counties invaded” metric, Morin and colleagues found that dogwood anthracnose had invaded 609 counties in the East (and additional areas in the West); the emerald ash borer had invaded 479 counties at the time of analysis; the hemlock woolly adelged had invaded 432 counties. Using the number of hosts impacted measure, oak wilt (Bretziella fagacearum) affected the second largest number of hosts – 61 (Potter et al. 2019a). [All these pests are described briefly here.]
Project CAPTURE (Potter and colleagues 2019a) evaluated 339 serious pests threatening one or more of 419 native tree species in the continental US. They included both native and introduced pests. They analyzed 1,378 pest-host combinations. They found that:
54% of the host tree species (228) are infested by an exotic pest – although only 28% of the 1,378 host/agent combinations involved pests are known to be non-native in origin.
Exotic agents have, on average, considerably more severe impacts than native pests.
Non-native pests had greater average severity on angiosperms than on conifers. (As an earlier blog documented, Mech and colleagues have reached a similar – although tentative – conclusion.)
Their estimate of the threat posed by non-native pests to forests – especially for the East – is an underestimate because established pests could spread to additional vulnerable areas and there is a high likelihood that new pests will be introduced. The Southeast was consistently a “coldspot” – despite the near extirpation of one understory tree – redbay.
Potter et al. (2019a) ranked forest threats in two ways. Four host families were at highest risk to alien pests, as measured by both the numbers of tree species affected and by the most host/agent combinations: Fagaceae (oaks, tanoaks, chestnuts, beech); Pinaceae (pines); Sapindaceae (soapberry family; includes maples and buckeye); Salicaceae (willows, poplars, aspens). When host families were ranked by the severity of the host/pest threat, Fagaceae was still at greatest risk, and Sapindaceae was still in the top four; however, Ulmaceae (elms) and Oleaceae (includes Fraxinus) replaced pines and willows.
A very interesting study was published by scientists based in the Blue Ridge Mountains of Virginia (Anderson-Teixeira et al. 2020). They contend that their area is a good example of what is happening more broadly in the Mid-Atlantic region.
Anderson-Teixeira et al. (2020) found that non-native pests have substantially impacted at least 24% of the 33 tree genera (eight genera) recorded as present in their study plots. They estimated that over the century beginning with the appearance of chestnut blight in the region and ending with the expected extirpation of ash trees, net live aboveground biomass (AGB) loss among affected species totaled roughly 6.6–10 kg m -2. Forty to sixty percent of this loss started before the Park initiated quantitative surveys of permanent plots in 1987. The authors estimated that chestnut contributed up to 50% of estimated AGB losses over the century. Consequently, the estimate has very high uncertainty.
Despite these losses, Anderson-Teixeira et al. (2020) found that both total aboveground biomass and diversity within individual study plots had largely recovered through increases in non-vulnerable genera.
Average above ground biomass across the plots established in Shenandoah National Park increased as the forest recovers from logging, farming, and other disturbances before formation of the Park. These increases were due primarily to reproduction and growth of tulip poplar (Liriodendron tulipifera) and growth (but not reproduction) of oaks. Net AGB biomass was lost in oak- and hemlock-dominated plots. At plots established in the neighboring Smithsonian Conservation Biology Institute, pests had caused relatively minor impacts on AGB.
Diversity of tree species also did not change much. In the Park, the average number of genera per plot declined only 3% between 1991 and 2013. Diversity at the landscape scale increased by two genera – from 26 to 28. Many individual plots, though, lost three genera due to non-native pests – chestnut, redbud, and hemlock. A fourth genus was lost due to stochastic change. At the same time, the plots gained six native genera). This finding might be skewed by the short duration of the study period, which missed initial declines in several taxa and captured only the initial stages of decline in ash.
Several taxa were lost from the monitoring plots but were not completely extirpated from the region. Even those species not “lost” suffered elevated mortality rates and steep declines in abundance and above-ground biomass. These declines have not been reversed. The exception was some oaks, which regained above ground biomass, but not abundance, following the gypsy moth outbreak in the 1980s and early 1990s.
Fei et al. (2019) found that losses in biomass due to non-native pests – as measured by FIA plot data – was greatest for ashes, elms, beech trees, and hemlocks..
Morin and colleagues found annual mortality rates had increased three-fold above background levels for ash, beech, and hemlock. They also calculated the present mortality rates for several species for which the majority of loss occurred before their study (consequently, they could not calculate a pre-invasion “background” rate to which present rates could be compared). These included American chestnut (mortality rate of 7%), butternut (mortality rate of 5.6%), and elm trees (mortality rate of 3.5%).
The CAPTURE Project (Potter et al. 2019a) identified fifteen host-agent combinations with the highest severity. Ten of these species are found in the Mid-Atlantic region:
American chestnut (Castanea dentata)
Allegheny chinquapin (C. pumila)
Carolina ash (Fraxinus caroliniana) ,
pumpkin ash (F. profunda)
Carolina hemlock (Tsuga caroliniana)
butternut (Juglans cinerea)
eastern hemlock (Tsuga canadensis)
white ash (Fraxinus americana)
black ash (F. nigra)
green ash (F. pennsylvanica)
Four of these species are in genera included among the eight genera evaluated in the study conducted in the Blue Ridge (Anderson-Teixeira et al. 2020): American chestnut, butternut, eastern hemlock, green and white ash. The four other genera in the Blue Ridge study were elm (Ulmus), oak (Quercus), redbud Cercis, and dogwood (Cornus). All except redbud are recognized by other sources as heavily affected by non-native pests – confirming Anderson-Teixeira et al. (2020)’s conclusion that findings on the Blue Ridge reflect the wider situation.
Anderson-Teixeira et al. (2020) note that several of these tree species have been declared imperiled by the International Conservation Union (IUCN): American chestnut, butternut, American elm, eastern hemlock, and ash species.
Anderson-Teixeira et al. (2020) report data on three taxa previously important in the canopy of Blue Ridge forests – chestnut, elms, and butternut. Chestnuts larger than 10 cm DBH had disappeared from the future site of Shenandoah National Park by 1910. Short-lived sprouts continue to be present in plots in the low-elevation Smithsonian Conservation Biology Institute. Two elm species were described as ‘‘sparse’’ in the 1939 qualitative survey. Elms have persisted at low densities, low biomass, and increasingly small sizes. Butternut was ‘‘common’’ in 1939, but had disappeared from Shenandoah NP by 1987. On the Smithsonian’s property, butternut declined from four living individuals in 2008 to two in 2018. The near disappearance of butternut reflects the national picture: FIA data show the species has decreased about 58% across its U.S. range since the 1980s – which is decades after butternut canker started having a detectable impact in the Midwest.
In the Park, oak-dominated plots lost on average 24.9% of individuals and 15% of aboveground biomass. After 1995, when the gypsy moth was better controlled by spraying of Bacillus thuringiensis var. curstaki, oak aboveground biomass increased gradually, driven by individual tree growth rather than new recruitment. Continued declines in oak abundance are attributable to oak decline and management actions (or inactions) that do not promote regeneration.
In a separate study, a group of oak experts went through a process of queries to identify the greatest threat to oaks now and in the future (Conrad et al. 2020). They initially identified the following threats as most important currently (descending order): gypsy moth, oak wilt, oak decline, climate change, and drought. The top five future threats were initially identified as climate change, oak wilt, sudden oak death, oak decline, and some unknown new or emerging (non-native) pest or pathogen. By the third round, after the experts thought about their colleagues’ responses, oak decline had replaced gypsy moth as the most critical threat currently. Attack by an unknown new or emerging (non-native) pest or pathogen replaced climate change as the most critical future threat. While there was not a complete consensus, the consensus was stronger on the threat from a new pest.
remnant eastern hemlock at Linderlost, Shenandoah National Park photo by F.T. Campbell
Anderson-Teixeira et al. (2020) reported that eastern hemlock was initially present in ten of Shenandoah plots, but was no longer recorded in the survey plots after 2007. (More than 20,000 insecticide-treated trees remain alive throughout Shenandoah NP).
Before arrival of the emerald ash borer, ash aboveground biomass was increasing in Shenandoah NP and stable on the Smithsonian Institute. EAB-caused mortality was first detected at the Smithsonian site in 2016 and accelerated steeply thereafter, exceeding 12.5% year by 2018. As of 2019, ash had lost 28% of individuals and 30% of aboveground biomass relative to 2016. Ninety-five percent of remaining live trees were considered “unhealthy’’ (Anderson-Teixeira et al. 2020).
eastern (flowering) dogwood; photo by F.T. Campbell
Unlike many studies, the Shenandoah study included understory species. Flowering dogwood declined by up to 90% from plots on the Smithsonian property; 2008–2019 mortality rates averaged 7.1%. Redbud declined by up to 76% from 1995 to 2018. The 2008–2019 mortality rates averaged 6.2% year.
Anderson-Teixeira et al. (2020) concede difficulty in estimating mortality due to less virulent or lethal pathogens, including Neofusicoccum spp. on redbud and Dutch elm disease on slippery elm.
Nevertheless, they believe their analysis probably underestimates the overall pest impacts because they did not analyze several other pest/host combinations known to be present in the Park: balsam woolly adelgid (Adelges piceae) on high-elevation populations of Abies balsamea; white pine blister rust (Cronartium ribicola) on eastern white pine (Pinus strobus); beech bark disease (Neonectria spp.) on American beech (Fagus grandifolia); thousand canker disease on walnut and butternut; and emerald ash borer on the novel host fringetree Chionanthus virginicus.
Another possible threat to oaks, winter moth (Operophtera brumata), is apparently now being controlled by the biocontrol agent Cyzenis albicans.
I am uncertain about the current status of two Diplodia fungi – Diplodia corticola and D. quercivora – link to blog which have been detected in both Florida and California. In Florida, almost all the symptomatic trees grow in cultivated settings where they are exposed to various stresses (Mullerin and Smith 2015).
However, host range studies indicate that 33 species of oaks and one species of chestnut that grow in the Southeast are vulnerable, to varying degrees, to D. corticola. Oaks in the red oak group (Section Lobatae) are more vulnerable than are white oaks (Section Quercus) (Mullerin and Smith 2015). In the test, the most vulnerable appear to be the following species native to the Southeast: Q. laurifolia, Q. virginiana, Q. geminata, Q. chapmanni, Q. laevis (turkey oak), Q. phellos, Q. pumila, and Q. incana (Dreaden et al. 2016).
What should we do?
Fei et al. (2019) noted that the losses to biomass would be exacerbated by the likely introduction of additional pests. They did not recommend any prevention actions.
Conrad et al. (2020) said their findings “lend support to national regulatory and awareness efforts to prevent the introduction and establishment of novel exotic insects and pathogens.”
Anderson-Teixeira et al. (2020) join others in declaring that future survival of the IUCN-listed species probably depends on conservation and restoration actions. They cite several sources, but not the CAPTURE Project – although the two studies reinforce each other. They specifically mention limiting invasive species’ spread through strengthened regulations and “enhanced plant biosecurity cyberinfrastructure”.
This last recommendation reinforces the message of Bonello et al. (2019) link to publication. We called for creation of a federal Center for Forest Pest Control and Prevention to implement end-to-end responses to forest pest invasions. One focus would be correcting the currently-inadequate focus on detection, development and deployment of genetic resistance while using modern techniques that allow for much faster breeding cycles.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
SOURCES
Anderson-Teixeira, K.J., V. Herrmann, W.B. Cass, A.B. Williams, S.J. Paull, E.B. Gonzalez-Akre, R. Helcoski, A.J. Tepley, N.A. Bourg, C.T. Cosma, A.E. Ferson, C. Kittle, V. Meakem, I.R. McGregor, M. N. Prestipino, M.K. Scott, A.R. Terrell, A. Alonso, F. Dallmeier, and W.J. McShea. Date? Long-Term Impacts of Invasive Insects and Pathogens on Composition, Biomass, and Diversity of Forests in Virginia’s Blue Ridge Mountains. Ecosystems
Bonello, P. , F.T. Campbell, D. Cipollini, A.O. Conrad, C. Farinas, K.J.K. Gandhi, F.P. Hain, D. Parry, D.N. Showalter, C. Villari, and K.F. Wallin. 2019. Invasive tree pests devastate ecosystems – A proposed new response framework. Frontiers
Conrad, A.O., E.V. Crocker, X. Li, W.R. Thomas, T.O. Ochuodho, T.P. Holmes, and C. D. Nelson. 2020. Threats to Oaks in the Eastern US: Perceptions and Expectations of Experts. Journal of Forestry, 2020, 14–27
Dreaden, Black, Mullerin, and Smith. Poster presented at the 2016 USDA Invasive Species Research Forum
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. Liebhold. 2019. Biomass losses resulting from insect and disease invasions in United States forests. Proceedings of the National academy of Sciences.
Guo, Q., S. Feib, K.M. Potter, A.M. Liebhold, and J. Wenf. 2019. Tree diversity regulates forest pest invasion. PNAS. www.pnas.org/cgi/doi/10.1073/pnas.1821039116
Morin, R.S., K.W. Gottschalk, M.E. Ostry, A.M. Liebhold. 2018. Regional patterns of declining butternut (Juglans cinerea L.) suggest site characteristics for restoration. Ecology and Evolution.2018;8:546-559
Morin, R. A. Liebhold, S. Pugh, and S. Fie. 2019. Current Status of Hosts and Future Risk of EAB Across the Range of Ash: Online Tools for Broad-Scale Impact Assessment. Presentation at the 81st Northeastern Forest Pest Council, West Chester, PA, March 14, 2019
Mullerin, S. & J.A. Smith. 2015. Bot Canker of Oak in FL Caused by Diplodia corticola & D. quercivora. Emergent Pathogens on Oak and Grapevine in North America. FOR318
Potter, K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019a. Important Insect and Disease Threats to United States Tree Species and Geographic Patterns of Their Potential Impacts. Forests. 2019 10 304.
Potter, K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019b. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation. (2019)
Dream Lake, Rocky Mountain National Park, with limber pine photo by F.T. Campbell
In this blog, I summarize two pest threats to the unique ecosystems on high-elevation mountain ridges in the West. At risk are several keystone tree species: the five-needle pines growing at high elevations (“high-five” pines) and subalpine fir. The invasive species causing this damage – white pine blister rust (WPBR; Cronartiumribicola) and balsam woolly adelgid (BWA; Adelges piceae) – are two of the most widespread non-native species threatening North American trees and affecting the highest proportion of host volumes (Morin).
The pines being killed by white pine blister rust are whitebark pine (Pinus albicaulis), limber pine (P. flexilis), Rocky Mountain bristlecone pine (P. aristata), foxtail pine (P. balfouriana), and southwestern white pine (P. flexilis var. reflexa). As of 2010, infestations had not been reported on Great Basin bristlecone pine (P. longaeva) and the Mexican white pine species. [Unless otherwise indicated, information on white pine blister rust is from a comprehensive review and synthesis published in the August 2010 issue of Forest Pathology (Vol. 40:3-4).]
As noted above, sub-alpine fir (Abies lasiocarpa) is also being affected – although less uniformly than the pines – by the balsam woolly adelgid.
Both of these pests arrived approximately a century ago, but they are still spreading and causing additional damage. White pine blister rust had spread widely throughout the West within 40 years of its introduction. Meanwhile, BWA spread among lowland and subalpine firs along the Pacific coast from California to British Columbia within 30 years of its first detection. Its spread eastward was slower, but relentless. It reached Idaho, Montana, Utah and interior British Columbia within 50 years. Also, BWA reached Alaska within 90 years of its introduction in California. These pests are perfect examples of how invasive species introduced long ago are dreaded “gifts that keep on giving”.
For a detailed discussion of these pests’ impacts, see the descriptions posted here. To summarize, though, WPBR is present in the ranges of eight of the nine vulnerable western white pines and has caused severe mortality to some species (Sniezko et. al. 2011). For example, 88% of the limber pine range in Alberta is affected (Dawe et al. 2020). WPBR is generally causing more damage to its hosts’ northern populations. Impact of the BWA are more subtle than WPBR. Also, impacts’ severity is linked to climatic conditions. For example, measurable decline on the Olympic Peninsula was greater on south-facing slopes. However, the study did not determine whether this reflected heat-loading and tree stress or more abundant subalpine fir on these slopes. An estimated 19-53% (average 37%) of subalpine fir trees had died on sample plots on one ridge over the 19 years since BWA was first detected there. Overall forest growth after 2007 could indicate partial recovery, a momentary pause in BWA invasion, or tree growth after severe weather events (Hutton 2015).
Ranges of Trees at Risk
Many of the host trees of these two pests are widespread; others are more narrowly endemic.
Limber pine reaches from Alberta and British Columbia south to mountain peaks in Arizona and New Mexico. Whitebark pine is found from Alberta and British Columbia to California and Nevada (USDA Plants database. Subalpine fir stretches from southeast Alaska along the Canadian Rockies coast into Washington, Oregon, east into Idaho, Montana, Wyoming, Colorado, Utah, even into scattered mountain ranges of Nevada and New Mexico (Hutton 2015).
Limber pine and subalpine fir are also found in a wide range of ecosystems within these ranges. Limber pine is found at both upper and lower tree lines in grassy, open forests; on exposed rocky slopes; and in dense, mixed-conifer stands. Subalpine fir is a pioneer species on ridges, alpine meadows, avalanche chutes, and lava beds (Ragenovich and Mitchell, 2006).
Before arrival of non-native pests or pathogens, these tree species have persisted for thousands of years under harsh conditions (Hutton 2015). Many of the individual trees were long-lived; some five-needle pines, e.g., bristlecone pines, have famously live for thousands of years. Core studies demonstrated that subalpine firs trees could live 272 years in the forests of Olympic National Park and 240 years in Glacier National Park (Hutton 2015). Surely loss of these trees – or even their conversion from large and old to small and short-lived – will result in significant destruction of these unique biomes.
All these trees play important roles in high altitude, unique ecosystems (Pederson et al. no date; Dawe 2020; Hutton 2015):
They retain ground water, slow the rate of snow melt, and maintain stream flow characteristics and water quality;
They curtail soil erosion and maintain slope stability; and
They provide high-value food and shelter to wildlife.
Whitebark and limber pines are famous for providing critical food for many wildlife species at high elevations —notably bears and nutcrackers (Compendium and Dawe 2020).
More Pest Threats
Other diseases, insects, and disturbances also pose serious threats to these tree species. The threats vary by region and age of the stand. They include – for the pines — mountain pine beetle (Dendroctonus ponderosae), dwarf mistletoe (Arceuthobium spp.), and various shoot, cone or foliage insects and pathogens. For subalpine fir, threats include western balsam bark beetle (Dryocoetes confusus), fir engraver (Scolytus ventralis), and the fir root bark beetle (Pseudohylesinus granulatus) (Hutton 2015). Trees are also damaged by bear and deer, seed predation by squirrels, wildfire, and biotic succession.
On Washington’s Olympic Peninsula, BWA initiates or predisposes subalpine fir for a novel disturbance complex. BWA-caused stress makes the trees more susceptible to moisture stress and endemic bark beetle attack. Surviving trees are subsequently subject to toppling by wind. A tree can die in a few years, survive with insects for up to 20 years, or recover, depending on duration, severity, and location of infestation, and local environmental conditions (Hutton 2015).
BWA study plots in the Cascade Range experienced subalpine fir mortality ranging from 7 to 79% (measured as stem counts, not basal area) over a 19 to 38 years study period. Higher mortality occurred at low-elevation, mesic sites. One stand experienced 40% mortality in 19 years, but lost the remaining 60% during a subsequent spruce budworm infestation. Most plots continued to show sporadic signs of adelgid presence and continued tree mortality. However, 41-69% of trees survived stem infestations (Hutton 2015).
How to Protect These Ecosystems
The seeds of both whitebark and limber pines are dispersed to newly disturbed, open areas by Clark’s nutcracker (Nucifraga columbiana). Furthermore, whitebark cones open to release seeds only after fire. This had led to expectations that prescribed fire could promote regeneration of these species. However, studies by Dawe (2020) and other have found that nutcracker seed caching behavior and seedling establishment are complex. Fire management might have to vary among regions, demanding consideration of stand characteristics,like openness and the presence of other tree species. For example, in the Colorado Front Range, limber pine can be replaced by subalpine fir when fire-free intervals are long. On the other hand, in Alberta, fire appeared to boost regeneration of the dominant tree species in the stands pre-fire. In the study areas, these were white spruce (Picea glauca) and lodgepole pine (Pinus contorta) (Dawe 2020). Dawe recommends protecting existing stands of limber pine through fire mitigation efforts, e.g., thinning and other fuel treatments, and supplementary planting of seedlings.
Efforts to find biocontrol agents to target the balsam woolly adelgid began in 1957; the original focus was on the insects’ damage to Fraser fir (Abies fraseri) in the southern Appalachians. More than 25 predatory species have been introduced from Europe and Asia. There was simultaneous research on native predators. None has had an impact on BWA populations in either the East or the West.
Neither white pine blister rust nor balsam woolly adelgid is considered a quarantine pest by federal officials, so there is no attempt to prevent their movement via interstate trade in Christmas trees, timber, or nursery stock. Hutton (2015) hypothesizes that the absence of regulatory measures targetting BWA arises from the pest’s gradual effect and the hosts’ not being commercially important as timber species (although several firs are important in horticulture and as Christmas trees). I think another factor is that the pests were introduced so long ago and are now widespread.
Efforts are under way to detect resistant genotypes to be used in breeding programs. Several of the lower-elevation five-needle pines vulnerable to WPBR have benefitted from extensive breeding efforts Whitebark pine has more recently been added to programs.
The eastern Fraser fir is the target of breeding – primarily for Christmas trees (APS). However, at least small-scale volunteer efforts have been carried forward by the Alliance for Saving Threatened Forests.
Hutton (2015) expresses hope that evolutionary pressure by BWA might enhance survival of more resistant forms of subalpine fir and lead to their gradual takeover. However, I ask, why leave it to chance?
In this context, I remind you of my involvement with a group (see Bonello et al. 2019) proposing creation of a federal Center for Forest Pest Control and Prevention to implement end-to-end responses to forest pest invasions – including overcoming the currently inadequate focus on detection, development and deployment of genetic resistance using modern techniques that allow for much faster breeding cycles.
I am puzzled that the Project CAPTURE places whitebark pine and subalpine fir only in Class A4.2, not among the highest priority species (Potter et al. 2019). As I blogged last spring, Project CAPTURE is part of a multi-partner effort to categorize and prioritize US tree species for conservation actions based on the threats and the trees’ ability to adapt to those threats. I find it puzzling because I am not sure I agree that these two species have a moderately high mean pest severity score – as required by the category. I am less puzzled by the assignment of a low adaptive capacity score.
Limber pine apparently ranks even lower in the Project CAPTURE priority process.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
SOURCES
A comprehensive review and synthesis of the history, ecology, and management of white pines threatened by white pine blister rust see the August 2010 issue of Forest Pathology (Vol. 40:3-4).
Bonello, P. , F.T. Campbell, D. Cipollini, A.O. Conrad, C. Farinas, K.J.K. Gandhi, F.P. Hain, D. Parry, D.N. Showalter, C. Villari, and K.F. Wallin. 2019. Invasive tree pests devastate ecosystems – A proposed new response framework. Frontiers
Dawe, D.A., V.S. Peters, M.D. Flannigan. 2020. Post-fire regeneration of endangered limber pine (Pinus flexilis) at the Northern extent of its range. Forest Ecology and Management 457 (2020) 117725
Hutton, K.M. 2015. A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy. University of Washington. Available here
Morin, R. Presentation to the 81st Northeastern Forest Pest Council Northeastern states forst agencies, Philadelphia, Pennsylvania, March 2019.
Potter, K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S. 2019. Prioritizing the conservation needs of US tree spp: Evaluating vulnerability to forest P&P threats, Global Ecology and Conservation (2019), doi: https://doi.org/10.1016/
Sniezko, R.A., M.F. Mahalovich, A.W. Schoettle, D.R. Vogler. 2011. Past and Current Investigations of the Genetic Resistance to Cronartium ribicola in High-elevation Five-needle Pines. In Keane, R.F., D.F. Tomback, M.P. Murray, and C.M Smith, eds. 2011. The future of high-elevation, five-needle white pines in Western North America. Proceedings of the High Five Symposium. 28-30 June, 2010. Missoula, MT.
eastern hemlocks in Great Smoky Mountains National Park
As
we all know, eastern (Tsuga canadensis)
and Carolina (T. caroliniana) hemlocks
have suffered huge losses due primarily to the introduced hemlock woolly
adelgid (Adelges tsugae – HWA). In New England, there has been more
than a 60% decrease in total hemlock basal area since 1997 and a virtual
absence of hemlock regeneration in HWA-infested areas. HWA continues to spread
– most recently into western Michigan and Nova Scotia (all information, unless
otherwise indicated, is from Kinahan et
al. 2020; full citation at end of this blog). [However, Morin and Liebhold
(2015) found that hemlock basal volume continued to increase for the first 20
years or so after invasion by the adelgid, due to ingrowth of immature
hemlocks. See “results” in Morin et al.,
full citation at the end of the blog.]
This
loss deprives us of a gorgeous tree … and unique habitats. Hemlock-dominated
forests were characterized by deep shade, acidic and slowly decomposing soil,
and a cool microclimate. They provided unique and critical habitat for many
terrestrial and aquatic species.
A
team of scientists based at the University of Rhode Island has carried out an
experiment comparing cuttings from eastern hemlocks apparently resistant to HWA
to susceptible ones. Matching sets of resistant and susceptible trees were
planted at eight sites in seven states – Ithaca and Bronx, NY; Boston; southern
CT; Lycoming County, PA; Thurmont, MD; southern WV; and Waynesville, NC. All plantings
were within or adjacent to forests containing HWA-infested hemlocks.
After
four years, 96% of the HWA-resistant hemlocks had survived, compared to 48% of
the control plants. The HWA-resistant plants were 32% taller, put out 18% more
lateral growth, had 20% longer drip lines, and were in 58% better condition.
HWA was found on trees at only three out of the eight plots. HWA density on
resistant eastern hemlocks was 35% lower than on HWA-susceptible hemlocks,
although this difference was not statistically significant.
Trees
in all eight plots were infested with elongate hemlock scale (Fiorinia externa – EHS), a second insect
damaging hemlocks in eastern North America. However, the HWA-resistant hemlocks
had EHS densities 60% lower than those of the controls.
Kinahan et al. note that identification and use of host tree populations’ potential for pest resistance has played a role in other programs managing non-native pests and pathogens, including Dutch elm disease and chestnut blight.
The
same scientists note that significant effort has been put into biocontrol or
insecticides for management of hemlock woolly adelgid, but without achieving the
desired improvement of forest health. Attempts to cross eastern hemlocks with
HWA-resistant hemlocks unfortunately produced no viable offspring. However,
Kinahan et al. were inspired to
explore possible genetic resistance within natural populations of eastern
hemlocks by the 1) evidence of resistance in Asian and western hemlocks; 2) the
different foliar terpene profiles in those species; and 3) the presence of
apparently healthy mature hemlock trees growing in proximity to heavily
infested trees.
They asked forest managers and other concerned groups to help locate stands with trees that were mature and apparently completely healthy, were located within HWA-devastated hemlock stands, and had not been chemically treated. They chose a small stand of eastern hemlocks growing within the Walpack Fish and Wildlife Management Area in northern New Jersey. This stand was called the “Bulletproof Stand”. They evaluated HWA resistance in five of these trees, then chose two for propagation and planting in the test.
New Jersey’s “bullet-proof stand” on the left photo by Richard Casagrande
The
trees were planted in September 2015. Due to funding gaps, they were not
revisited for four years. Thus, Kinahan et
al. re-evaluated the resistant and vulnerable trees in Autumn 2019 – with
the results I reported above.
Does this study
prove that clonal propagation of apparently resistant hemlocks is an effective
strategy to restore the species?
It
is not that simple.
The
difference in survival and condition was striking, but the authors note several
caveats:
1)
they had not recorded pre-experiment data on plant height or other variables,
so they cannot be certain that variation in initial plant height or dripline
did not contribute to current treatment-level differences in these variables.
2)
they cannot distinguish between the impacts of HWA and EHS on plant growth.
3)
since they could not monitor the planting sites for four years, they cannot
definitively link increased mortality of HWA-susceptible trees to higher pest
densities. However, the lower pest densities and higher survival of
HWA-resistant hemlocks are consistent with herbivore-driven tree mortality.
They
also cannot assess the impact of other environmental stressors (drought, cold,
etc.) on their results.
4)
The small number of trees planted at each site prevented detailed site-level
analyses.
The
scientists conclude that their work is most appropriately viewed as a ‘proof of
concept’ experiment highlighting the need
for future research exploring how HWA-resistant eastern hemlocks might best be
integrated into existing HWA management.
Unfortunately, the
Rhode Island researchers report they cannot persuade the US Forest Service to
support continuing this effort. Will these promising hints not result in
action?
Kinahan
et al. stress the importance of the
reduced pest densities (both HWA and EHS) on the putatively resistant hemlocks.
They think this might be a result of the higher terpene concentrations in the
twigs and needles. Finally, they note that lower densities of sap-feeding
herbivores may also indirectly provide protection against other consumers,
including gypsy moth (Lymantria dispar)
and hemlock looper (Lambdina fiscellaria).
SOURCE
Kinahan,
I.G., G. Grandstaff, A. Russell, C.M. Rigsby, R.A. Casagrande, and E. L.
Preisser. 2020. A four-year, seven-state reforestation trial with eastern hemlocks
(Tsuga canadensis) resistant to hemlock woolly adelgid (Adelges tsugae).
Forests 11: 312
Morin, R.S. and A.M. Liebhold. 2015. Invasions by two non-native insects alter regional forest species composition and successional trajectories. Forest Ecology and Management 341 (2015).
Posted by Faith Campbell
P.S. I have been working with colleagues to promote a more coordinated and well-funded program to combat non-native forest pests – including much greater reliance on identifying and breeding resistance to the pest. Visit here to see this effort.
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
prickly pear cacti in Big Bend National Park photo by Blake Trester, National Park Service
The cacti that are such important components of desert ecosystems across nearly 2 million square miles straddling the U.S.-Mexico border are under threat from non-native insects – as I have noted in earlier blogs. Of course, cacti are important in other ecoregions, too – I wrote recently about the columnar cacti in the dry forests of Puerto Rico.
Flat-padded prickly pear cacti of the genus Opuntia are threatened by the cactus moth, Cactoblastis cactorum.
In 1989, the cactus moth was found
in southern Florida, to which it had spread from the Caribbean islands (Simonson
2005). Recently, the moth was found to have spread west as far as the Galveston,
Texas, area and near I-10 in Columbus, Texas, about 75 miles west of central
Houston (Stephen Hight, pers. com.) Two
small outbreaks on islands off Mexico’s Caribbean coast have been eradicated.
In Florida, the cactus moth has
caused considerable harm to six native species of prickly pear, three of which
are listed by the state as threatened or endangered.
When the cactus moth reaches the
more arid regions of Texas, it is likely to spread throughout the desert
Southwest and into Mexico. In the American southwest, 31 Opuntia species are at risk; nine of them are endemic, one is endangered.
Mexico is the center of endemism for the Opuntia
genus. In Mexico, 54 Opuntia species
are at risk, 38 of which are endemic (Varone et al. 2019; full citation at end of this blog).
The
long-term effects of the cactus moth on these North American Opuntia are unknown because there may be
substantial variations in tolerance. The attacks observed in the Caribbean
islands have shown great variability in various cactus species’ vulnerability (Varone et al. 2019).
The Opuntia cacti
support a diversity of pollinators as well as deer, javalina (peccaries),
tortoises, and lizards. Prickly pears also shelter packrats and nesting birds (which
in turn are fed on by raptors, coyotes, and snakes), and plant seedlings. Their
roots hold highly erodible soils in place (Simonson 2005).
While scientists have been concerned about the possible impacts of the cactus moth since it was detected in Florida 30 years ago, a substantial response began only 15 years later. The U.S. Department of Agriculture began trying to slow the spread of the cactus moth in 2005 (Mengoni Goñalons et al. 2014), with a focus on surveys and monitoring, host (cactus) removal, and release of sterile males. This program was successful at slowing the moth’s spread and eradicating small outbreaks on offshore islands of Alabama, Mississippi, and Mexico.
Cactus moth damage to native cacti in Florida photo by Christine Miller, UF/IFAS
However,
the moth continued to spread west and the program never received an appropriation from Congress. The primary funding source was a US – Mexico
Bi-National Invasive Cactus Moth Abatement Program. Both countries contributed
funds to support the research and operational program to slow the spread in the
U.S. Funds were provided through USDA Animal and Plant Health and Inspection
Service (APHIS) and the Mexican Secretariat of Agriculture, Livestock, Rural
Development, Fisheries and Food (SEGARPA). Unfortunately,
funding was reduced by both entities and became inadequate to maintain the
Bi-National Program.
Therefore, in 2012, APHIS abandoned
its regional program and shifted the focus to biocontrol. This is now
considered the only viable control measure in the desert Southwest where vulnerable
cacti are numerous and grow close together. The biocontrol project has been funded
since 2012 through the Plant Pest and Disease Management and Disaster
Prevention program (which receives funding through the Farm Bill). It has
received a total of slightly more than $2
million over seven years. More than half the funds went to the quarantine
facility to support efforts to rear non-target hosts and verify the biocontrol
agent’s host specificity. About a quarter of the funds supported complementary
work of an Argentine team (both the cactus moth and the most promising
biocontrol agent are native to Argentina). Much smaller amounts have supported
U.S.-based scientists who have studied other aspects of the cactus moth’s
behavior and collected and identified the U.S. moths being tested for their
possible vulnerability to attack by a biocontrol wasp.
Here are
details of what these dedicated scientists achieved in just the past seven
years at the relatively low cost of roughly $2 million. Unfortunately, the project now faces a funding crisis and
we need to ensure they have the resources to finish their work.
Some
Specifics of the BioControl Program
After literature reviews, extensive collections,
and studies in the cactus moth’s native habitat in Argentina (Varone et al. 2015), a newly described wasp, Apanteles
opuntiarum (Mengoni Goñalons et al. 2014), has been determined to be host
specific on Argentine Cactoblastis species and the most promising
candidate for biocontrol. Wasps were collected in Argentina and sent to
establish a colony in a quarantine facility in Florida to enable host
specificity studies on North American Lepidoptera (Varone et al. 2015).
Quarantine
host specificity studies and development of rearing technology has not been straightforward. Initially, it was
difficult to achieve a balanced male/female ratio in the laboratory-bred generations;
this balance is required to maintain stable quarantine laboratory colonies for
host range testing. This difficulty was overcome. A second challenge was high
mortality of the cactus-feeding insects collected in the Southwest that were to
be test for vulnerability to the biocontrol wasp. These desert-dwellers don’t
do well in the humid, air-conditioned climate of the quarantine facility! For
these difficult-to-rear native insects, scientists developed a molecular
genetics method to detect whether eggs or larvae of the cactus moth parasitoid were
present inside test caterpillars after they were exposed to the wasps. For easy
to rear test insects, caterpillars are exposed to the wasps and reared to
adulthood. Host specificity tests have been conducted on at least five species
of native U.S. cactus-feeding caterpillars and 11 species of non-cactus-feeding
caterpillars (Srivastava
et al. 2019; Hight pers.comm.).
To
date there has been no instance of
parasitism by Apanteles opuntiarum on either lepidopteran non-target species or
non-cactus-feeding insects in the Florida quarantine or in field collections in
Argentina (Srivastava et al.
2019; Varone et al. 2015; Hight pers.comm.).
The scientists expected to complete host-specificity testing in the coming months, then submit a petition to APHIS requesting the release of the wasp as a biocontrol agent. Unfortunately, the project’s request for about $250,000 in the current year was not funded. This money would have funded completion of the host specificity testing, preparation of a petition to APHIS in support of release of the biocontrol agent into the environment, and preparation of the release plan.
Meanwhile,
what can we expect regarding the probable efficacy of the anticipated biocontrol
program?
Some
of the wasp’s behavioral traits are encouraging. The wasp is widely present in
the range of the cactus moth, and persisted in these areas over the years of
the study. The wasp can deposit multiple eggs with each “sting”. Multiple wasps
can oviposit into each cactus moth without detriment to the wasp offspring. Unmated wasp females
produce male offspring only, whereas mated females produce mixed offspring
genders. In the field, female wasps attack cactus moth larvae in a variety of
scenarios: they wait at plant access holes to sting larvae when they come
outside to defecate; they attack larvae when they are moving on the surface of
the pads; they can sting the youngest cactus moth larvae through the thin plant
wall of mined the pads; and they enter large access holes created by older
larvae and attack larger larvae. The wasps are attracted by the frass
(excrement) left on the outside of the cactus pads by cactus moth larvae (Varone
et al. 2020).
However, I wonder about the extent
to which the cactus moth is controlled by parasitoids in Argentina. Cactoblastis eggs are killed
primarily by being dislodged during weather events (rain and wind) and by
predation by ants. First instar larvae are killed primarily by the native Argentine
cactus plants’ own defenses – thick cuticles and release of sticky mucilage when
the young larvae chew holes into the pads where they enter and feed internally.
As larvae feed and develop inside the pads, the primary cause of mortality is
natural enemies.
Of
all the parasitoid species that attack C. cactorum, A. opuntiarum
is the most abundant and important. When the larvae reach their final state (6th
instars), they leave the pads and find pupation sites in plant litter near the
base of the plants. It is at this stage that the parasitism from A.
opuntiarum is detected in the younger larvae that were attacked while
feeding inside pads. As the moth larva begins to spin silk into which to
pupate, larvae of the wasp erupt through the skin of the caterpillar and pupate
within the silk spun by the moth. Predation by generalists (ants, spiders,
predatory beetles) accounted for high mortality of the unprotected last instar
and pupae (Varone et al. 2019).
Finally,
the cactus moth has three generations per year when feeding on O. stricta in the subtropical and tropical coastal areas of the Americas
and the Caribbean. In Argentina, on its native host, the moth completes
only two generations per year (Varone et
al. 2019).
How to
Get the Program Support Needed
Opuntia in Big Bend National Park Photo by Cookie Ballou, National Park Service
To date, no organized
constituency has advocated for protection of our cacti from non-native insect pests.
Perhaps now that the Cactoblastis
moth is in Texas, the threat it represents to our desert ecosystems will become
real to conservationists and they will join the struggle. The first step is to
resolve the funding crisis so that the agencies can complete testing of the biocontrol
agent and gain approval for its release. So now there is “something people can
do” – and I hope they will step forward.
I hope Americans are not actually indifferent
to the threat that many cacti in our deserts will be killed by non-native
insects. Many are key components of the ecosystems within premier National
Parks, and other protected areas. Cacti also are beautiful treasures in
botanical gardens. I hope conservationists will agree that these threats must
be countered, and will help to ensure
funding of the final stages of the biocontrol tests.
Sources
Mengoni Goñalons, C., L. Varone, G. Logarzo, M. Guala, M.
Rodriguero, S.D. Hight, and J.E. Carpenter. 2014. Geographical range & lab
studies on Apanteles opuntiarum (hymenoptera: braconiDae) in AR, a candidate
for BC of Cactoblastis cactorum (Lepidoptera: Pyralidae) in North America. Florida
Entomologist 97(4) December 2014
Srivastava, M., P. Srivastava, R. Karan, A. Jeyaprakash,
L. Whilby, E. Rohrig, A.C. Howe, S.D. Hight,
and L. Varone. 2019. Molecular detection method developed to track the
koinobiont larval parasitoid Apanteles opuntiarum (Hymenoptera: Braconidae) imported from Argentina to control Cactoblastis cactorum (Lepidoptera:
Pyralidae). Florida Entomologist 102(2): 329-335.
Varone, L., C.M. Goñalons, A.C. Faltlhauser, M.E. Guala,
D. Wolaver, M. Srivastava, and S.D. Hight. 2020. Effect of rearing Cactoblastis cactorum on an artificial
diet on the behavior of Apanteles
opuntiarum. Applied Entomology DOI: 10.1111/jen.12731.
Varone,
L., G. Logarzo, J.J. Martínez, F. Navarro, J.E. Carpenter, and S.D. Hight.
2015. Field host range of Apanteles
opuntiarum (Hymenoptera: Braconidae) in Argentina, a potential biocontrol
agent of Cactoblastis cactorum
(Lepidoptera: Pyralidae) in North America. Florida Entomologist — Volume 98,
No. 2 803
Varone, L., M.B. Aguirre, E. Lobos, D. Ruiz Pérez, S.D. Hight, F. Palottini, M. Guala, G.A. Logarzo. 2019. Causes of mortality at different stages of Cactoblastis cactorum in the native range. BioControl (2019) 64:249–261
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
SOD in California; photo by Joseph O’Brien. courtesy of Bugwood
We know that the international trade in living plants is a major pathway by which tree-killing pathogens are being spread – some of them again and again. According to Grünwald etal. (2019), Phytophthora ramorum, the pathogen that causes Sudden Oak Death (SOD), has been introduced to North America and Europe – probably from Asia – at least five times. One lineage or genetic strain – EU1 – has been established on both continents (strains explained here). There is strong evidence of two separate introductions to Oregon, at least 12 to California.
Jung
et al. 2015 state definitively that
the international movement of infested nursery stock and planting of
reforestation stock from infested nurseries have been the main pathway of
introduction and establishment of Phytophthora
species in European forests.
Clive Btasier in Vietnam photo from UK Forest Research
Jung et al. 2020 have demonstrated that P. ramorum probably originated in
Vietnam. This region appears to be a center of
diversity for Phytophtoras and other
Oomycetes: baiting of soil and streams resulted in the detection of 13
described species, five informally designated taxa, and 21 previously unknown
taxa of Phytophthoras plus at least
15 species in other genera. Noting the risk associated with any trade in plants
from this region, the authors re-iterated past appeals that the international
phytosanitary system replace the “outdated and scientifically flawed
species-by-species regulation approach based on random visual inspections for
symptoms of described pests and pathogens” by instituting “a sophisticated
pathway regulation approach using pathway risk analyses, risk-based inspection
regimes and molecular high-throughput detection tools.”
Pathogen’s Spread Proves U.S.
Domestic Regulations Governing Nursery Trade Are Inadequate
Last year I blogged about the most recent spread of Phytophthora ramorum through the nursery trade. As of now, we know that shipments of potentially infected plants had been sent to 18 states. Infected stock had been detected in nurseries in seven of these (Iowa, Illinois, Indiana, Kansas, Missouri, Nebraska, Oklahoma) plus the source state, Washington [COMTF Newsletter August 2019].
Since then, I learned [COMTF newsletter for December 2019] that these plants were infected by the NA2 strain of the pathogen. This is the first time that this strain has been shipped to states outside the West Coast. It is unclear what the impact will be if – as is likely – infested plants are still extant in purchasers’ yards. Both the NA1 strain (the strain established in most infested forests of California and Oregon) and the NA2 strain belong primarily to the A2 mating type, so the potential spread of NA2 lineages might not exacerbate the probability of sexual reproduction of the pathogen.
I applaud agencies’ funding of
genetic studies to determine the lineage of the pathogen involved. It not only
helps narrow the possible sources of infected plants, but also could be
important in determining risk and management options.
I have long criticized USDA’s P. ramorum regulatory program – see Fading Forests III and my blogs discussing the most recent revisions to the regulations here and here. I believe that both the earlier regulations and the revisions finalized last May provide inadequate protection for America’s forests.
The updated regulations do take a couple of important positive steps. First, APHIS is now authorized to sample water, soil, pots, etc. – and to act when it finds evidence of the pathogen’s presence. APHIS also now mandated nurseries found to be infested to carry out a “critical control point analysis” to determine practices which facilitated establishment and persistence of P. ramorum.
However, these improvements are
severely undermined by continuing the five-year-old practice of limiting close
scrutiny to only those nurseries that tested positive for the pathogen in the
recent past. The flaw in this approach was starkly demonstrated by the
pathogen’s spread in 2019. The Washington State nursery that was the source of
the infected plants had not previously been positive, so it was under routine
nursery regulation, not the more stringent federal P. ramorum program.
Too often various iterations of the regulations have allowed infected plants to be shipped. Between 2003 and 2011, a total of 464 nurseries located in 27 states tested positive for the pathogen, the majority as a result of shipments traced from infested wholesalers (Campbell). The number of nurseries found to have infected plants has since declined, but not dropped to zero. These include 34 nurseries in 2010 (COMTF February 2011 newsletter), 21 in 2012, and 17 in 2013 (Pfister). During 2014, state inspectors detected the SOD pathogen in 19 nurseries – 11 in the three west-coast states and eight in other parts of the country (Maine-1, New York-2, Texas-1, and Virginia-4) COMTF newsletter December 2014). Despite the continuing presence of the pathogen in the nursery trade, APHIS formalized existing practices that narrowed the regulators’ focus to only those nurseries with a history of pathogen presence. This approach has been shown to fail – we need APHIS and the states to find a way to broaden their scrutiny.
The most immediate impact of the continuing presence of P. ramorum in the nursery trade is the burden borne by eastern states’ departments of agriculture. They are obligated to seek out in-state nurseries that might have received infected plants; inspect those plants; and destroy the infected plants, test nearby plants, and try to find and retrieve plants that had been sold. The heaviest, and most direct, burden is borne by the receiving nurseries. Anger about bearing this burden for 15 years doubtless prompted the National Plant Board to adopt a tart resolution calling on APHIS to carry out a review of its communications to the states during the 2019 incident. The NPB also questioned whether current program processes and guidance are effective in preventing spread of this pathogen.
Unfortunately, the NPB had not
commented formally on the rule change when it was proposed.
The states’ frustration is
exacerbated by the fact that under
the Plant Protection Act, when APHIS takes a regulatory action it prevents
states from adopting more stringent regulations. While the law allows for
exceptions if the state can demonstrate a special need, none of the five
applications for an exemption pertaining to P.
ramorum was approved (Porter and Robertson 2011). I have been unable to
find evidence of petitions submitted in the nine years since 2011.
In Case You Needed A
Reminder: P. ramorum is a Dangerous Pathogen
– as Proved by the Situation in the West states and Abroad
Continuing Intensification of the Already Bad Infestations in the West
tanoak mortality in Big Sur photo courtesy of Matteo Garbelotto, UC Berkeley
As of 2014 (see COMTF November 2018 newsletter available here), perhaps 50 million trees had been killed by P. ramorum in California and Oregon. The vast majority were tanoaks (Notholithocarpus densiflorus) – an ecologically important tree.
Since
2014, the disease has intensified and spread in
response to recent wet winters. In 2016 (see COMTF
November 2016 newsletter here) disease was detected for the first time in a fifteenth California county and new outbreaks or more severe infestations were recorded in seven other counties. In 2019, SOD was detected in the sixteenth county. Tanoak mortality in California increased by more than 1.6 million trees across 106,000 acres in 2018.
Perhaps more disturbing, the disease has also intensified on the eastern side of San Francisco Bay – an area thought to be less vulnerable because it is drier and where there are fewer of the principal sporulation host, California bay laurel (see COMTF March 2017 newsletter here).
A second disturbing event is the detection in Oregon forests of the EU1 strain of Phytophthora ramorum. The August 2015 detection was the first instance of this strain being detected in a forest in North America. Oregon authorities prioritized removing EU1-infected trees and treating (burning) the immediate area, which had expanded to more than 355 acres – all within the quarantine area in Curry County. The legislature provided $2.3 million for SOD treatments for 2017-2019 (Presentation by Chris Benemann of Oregon Department of Agriculture to the Continental Dialogue on Non-Native Forest Insects and Diseases; reported here).
The EU1 lineage is a different
mating type than the NA1 lineage already established in Oregon. Scientists
should study P. ramorum populations
in Vietnam and Japan, where both mating types are present, to determine whether
they are reproducing sexually. There is also the risk that the EU1 lineage
might be more aggressive on conifers – as it has been in the United Kingdom (Grünwald etal. 2019).
The EU1 infestation was introduced to the forest from a nursery. The nursery had carried out the APHIS-mandated Confirmed Nursery Protocol, then closed. I ask, what does this apparent transmission from nursery to forest say about the risk of transmission? Does it raise questions about the efficacy of the confirmed nursery protocol to clean up the area? Remember that a pond at the botanical garden in Kitsap, Washington has repeatedly tested positive, despite several applications of the clean-up protocols.
(For a discussion of the implications of mixing the various strains of P. ramorum, visit here)
These disasters remind us how sad it
is that California and federal officials did not adopt aggressive management
efforts aimed at slowing the pathogen’s spread at an early stage of the epidemic. Experts on modeling the
epidemiology of plant disease concluded three years ago that the sudden oak
death epidemic in California could have been slowed considerably if aggressive and
well-funded management actions had started in 2002 (Cunniffe, Cobb,
Meentemeyer, Rizzo, and Gilligan 2016).
The Oregon Department of Forestry commissioned a study of the economic impact of the P. ramorum infestation that found few economic impacts to date, but potentially significant impacts in the future. It also noted potential harms to tribal cultural values and the “existence value” of tanoak-dominated forests and associated obligate species.
Situation Abroad
The situation in Europe is even worse than in North America. Two strains of P. ramorum are widespread in European nurseries and in tree plantations and wild heathlands of western the United Kingdom and Ireland. and here and here. Jung et al. 2015 found 56 Phytophthora taxa in 66% of 2,525 forest and landscape planting sites across Europe that were probably introduced to those sites via nursery plantings.
larch plantation in UK killed by P. ramorum photo from UK Forest Research
In Australia, Phytophthora dieback has infected more than one million hectares in Western Australia. More than 40% of the native plant species of the region are vulnerable to the causal agent,P. cinnamomi
Barber
et al. 2013 reported 9 species of Phytophthora associated with a wide variety
of host species in urban streetscapes, parks, gardens, and remnant native
vegetation in urban settings in Western Australia. Phytophthora species were recovered from 30% of sampled sites.
In
New Zealand, the endemic – and huge, long-lived – kauri tree (Agathis australis) is also suffering
severe impacts from Phytophtoras and
other pathogens (Bradshaw et al.
2020)
See
the IUFRO Working Party 7.02.09 ‘Phytophthora Diseases of Forest Trees’ global
overview (Jung et al. 2018), which covers
13 outbreaks of Phytophthora-caused
disease in forests and natural ecosystems of Europe, Australia and the
Americas.
The situation in
the Eastern United States is Unclear
After 15 years of the nursery trade carrying P. ramorum to nurseries – and possibly yards and other plantings – in states east of the 100th Meridian, what is the risk that these forests will become infested? No one knows. We do known that the pathogen has been detected from 11 streams in six eastern states – four in Alabama; one in Florida; two in Georgia; one in Mississippi; one in North Carolina; and two in Texas. P. ramorum has been found multiple times in eight of these streams – two steams in Alabama, one each in Mississippi and North Carolina (see COMTF April 2019 newsletter available here). While established vegetative infections have not been detected, the question remains: how is the pathogen persisting? Scientists agree that P. ramorum cannot persist in the water; it must be established on some plant parts (roots?) or in the soil. Still, Grünwald etal. (2019) report that there is little evidence of plant infections resulting from stream splash in Oregon.
Unfortunately, fewer states are participating in the stream surveys – which are operated by the USDA Forest Service. In 2010, 14 states participated; in 2018, only seven (Alabama, Georgia, Mississippi, North Carolina, Pennsylvania, South Carolina, and Texas). Florida and Tennessee recently dropped out. The number of streams surveyed annually also has dropped – from 95 at the highest to only 47 in 2018 (see COMTF April 2019 newsletter available here). This reduced scrutiny makes it less likely that any infestation on plants will be detected. Risk maps (reproduced in Chapter 5 of Fading Forests III here) developed over more than a decade indicate that forests in the southern Appalachians and Ozarks are vulnerable to SOD.
Risks to other
plants
The risk from Phytophthoras is not just P. ramorum and trees! Swiecki et al. 2018 report a large and increasingly diverse suite of introduced Phytophthora species pose an ever greater threat to both urban and non-urban plant communities in California. These threats are linked to planting of nursery stock. See also the information posted here.
Jung et al. 2018 cite numerous other authors’ findings of multiple Phytophthoras in Oregon and. California nurseries as well as in nurseries in various eastern states.
Nor is Phytophthoras the only pathogenic genus to pose a serious risk to America’s trees. I remind you of the fungus Fusariumeuwallacea associated with the Kuroshio and polyphagous shot hole borers, which is known to kill at least 18 species of native plants in California and additional species in South Africa. The laurel wilt fungus kills many trees and shrubs in the Lauraceae family. ‘Ohi‘a or myrtle rust kills several shrubs native to Hawai`i and threatens a wide range of plants in the Myrtaceae family in Australia and New Zealand; rapid ‘ohi‘a death fungi (Ceratocystis huliohia and Ceratocystis lukuohia) [All described here] are killing the most widespread tree on the Hawaiian Islands.
Solutions – complete &
implement modernized international and domestic phytosanitary regulations
Clearly,
standard phytosanitary practice of regulating pests known to pose a threat
does not work when many – if not most – of the damaging pests are unknown to
science until introduced to a naïve ecosystem where they start causing
noticeable levels of damage. We need a more proactive approach – as has long been
advocated by forest pathologists, including Clive Brasier 2008 and later,
Santini et al. 2013, Jung et al. 2016, Eschen et al. 2017.
National and international phytosanitary agencies have taken some steps toward adopting policies and programs that all hope will be more effective in preventing the continued spread of these highly damaging tree-killing pests. First, APHIS has had authority since 2011 – through the Not Authorized for Importation Pending Pest Risk Assessment (NAPPRA) program — to prohibit temporarily imports of plants suspected of transporting known damaging pathogens until the agency has conducted a pest risk analysis. However, utilization has lagged: only three sets of species have been proposed for listing in NAPPRA in the eight and a half years since the program was instituted in 2011. The third list of proposed species is currently open for public comment.
Another
weakness is that the program still focuses on organisms known to pose a risk.
Second, in 2018 APHIS completed a decades-long effort to revise its plant import regulations (the “Q-37” regulations). APHIS now has authority to require foreign suppliers of living plants to carry out “hazard analysis and critical control point” programs and adopt integrated pest management strategies to ensure that the plants are pest-free during production and transport.
However, implementation of this new
authority depends on APHIS negotiating agreements with individual countries
that would govern specific types of plants exported to the U.S. APHIS has not
yet announced completion of any programs under this authority. Nor is it clear
which taxa or countries APHIS will prioritize.
APHIS’ action was anticipated by the international plant health community. In 2012, member states in the International Plant Protection Convention adopted International Standard for Phytosanitary Measure 36 (ISPM#36) The standard sets up a two-level system of integrated measures, which are to be applied depending on the pest risk identified through pest risk analysis or a similar process. The “general” integrated measures are widely applicable to all imported plants for planting. The second level includes additional elements designed to address higher pest-risk situations that have been identified through pest risk analysis or other similar processes.
However,
the preponderance of international efforts to protect plant health continues to
rely on visual inspections that look for species on a list of those known to be
harmful. Yet we know that most damaging Phytophthoras
were unknown before their introduction to naïve ecosystems.
Furthermore,
use of fungicides and fungistatic chemicals – that mask infections but do not
kill that pathogen – is still allowed before shipment.
(For more complete analyses of the Q-37 revision and ISPM#36, see chapters five and four, respectively, of Fading Forests III.)
The
nursery industry is working with state regulators and APHIS to develop a voluntary
program utilizing integrated measures –
the Systems Approach to Nursery Certification (SANC) program. https://sanc.nationalplantboard.org/
SOURCES
Bradshaw
et al. 2020. Phytophthora
agathidicida: research progress, cultural perspectives and knowledge gaps in
the control and management of kauri dieback in New Zealand. Plant Pathology
(2020) 69, 3–16 Doi: 10.1111/ppa.13104
Brasier
CM. 2008. The biosecurity threat to the UK and global environment from
international trade in plants. Plant Pathology 57: 792–808.
Brasier, C.M, S. Franceschini, A.M.
Vettraino, E.M. Hansen, S. Green, C. Robin, J.F. Webber, and A.Vannini. 2012.
Four phenotypically and phylogenetically distinct lineages in Phytophthora lateralis
Fungal Biology. Volume 116, Issue
12, December 2012, Pages 1232–1249
Campbell, F.T.
Calculation by F.T. Campbell from tables in U.S. Department of Agriculture,
Animal and Plant Health Inspection Service – National Plant Board. 2011. Phytophthora ramorum Regulatory Working
Group Reports. January 2011.
Cunniffe, N.J., R.C. Cobb, R.K.
Meentemeyer, D.M. Rizzo, and C.A. Gilligan. Modeling
when, where, and how to manage a forest epidemic, motivated by SOD in Calif. PNAS, May 2016 DOI: 10.1073/pnas.1602153113
Grünwald,
N.J., J.M. LeBoldus, and R.C. Hamelin. 2019. Ecology and Evolution of the
Sudden Oak Death Pathogen Phytophthora ramorum. Annual Review of Phytopathology
date? #?
Jung
T, Orlikowski L, Henricot B, et al.
2016. Widespread Phytophthora infestations in European nurseries put forest,
semi-natural and horticultural ecosystems at high risk of Phytophthora
diseases. Forest Pathology 46: 134–163.
Jung,
T., A. Pérez-Sierra, A. Durán, M. Horta Jung, Y. Balci, B. Scanu. 2018. Canker
and decline diseases caused by soil- and airborne Phytophthora species in
forests and woodlands. Persoonia 40, 2018: 182–220 Open Access!
Jung,
T. et al. 2015. Widespread Phytophthora infestations in European
nurseries put forest, semi-natural and horticultural ecosystems at high risk of
Phytophthora disease. Forest
Pathology. November 2015; available from Resource Gate
Knaus, B.J., V.J. Fieland, N.J.
Grunwald. 2015. Diversity of Foliar Phytophthora
Species on Rhododendron in Oregon
Nurseries. Plant Disease Vol 99, No. 10 326 – 1332
Pfister,
S. USDA APHIS. Presentation to the National Plant Board, August 2013
Porter, R.D. and N.C.
Robertson. 2011. Tracking Implementation of the Special Need Request Process
Under the Plant Protection Act. Environmental Law Reporter. 41.
Santini
A, Ghelardini L, De Pace C, et al.
2013. Biogeographic patterns and determinants of invasion by alien forest
pathogens in Europe. New Phytologist 197: 238–250.
Swiecki,
T.J., E.A. Bernhardt, and S.J. Frankel. 2018. Phytophthora root disease and the need for clean nursery stock in
urban forests: Part 1 Phytophthora
invasions in the urban forest & beyond. Western Arborist Fall 2018
Tsao PH.
1990. Why many Phytophthora root rots and crown rots of tree and horticultural
crops remain undetected
wiliwili flower photo by Forrest and Kim Starr, courtesy of creative commons
Hawaii’s
dryland forest is a highly endangered ecosystem. More than 90% of dry forests
are already lost due to habitat destruction and the spread of invasive plant
and animal species. However, a new publication documents some recovery of
wiliwili trees from one major pest. At the same time, a new pest is spreading
and killing naio, a critical dryland shrub. Both pests originated in countries that have rarely
if ever been a source of U.S. pests. This is worrying because phytosanitary
agencies have their hands full with imports from the usual sources. The role of
California as a source of invasive species in Hawai`i has long deserved federal
attention – but as far as I know has not received it.
Hope for Wiliwili Trees
The
Hawaiian endemic wiliwili tree, Erythrina
sandwicensis, occurs in lowland dry forests on all the major islands from
sea level to 600 m. Wililwili is a dominant overstory tree in these habitats.
(Unless otherwise noted, the principal source is Kaufman et al. in press – full
citation at end of blog.)
The tree has been severely affected by the introduced Erythrina gall wasp, Quadrastichus erythrinae (EGW). The gall wasp was detected on Oahu in 2005 and quickly spread to the other Hawaiian islands.
Arrival
of the EGW on Oahu was part of the insect’s rapid global range expansion. Originally from East Africa, it was first
detected in the Mascarene Islands and Singapore in 2003. At the time, it was unknown
to science. Within a few years it had spread across Asia, many Pacific islands
(including Hawai`i), and to the Americas, including Florida in 2006, Brazil in 2014
(Culik 2014), and Mexico in 2017 (Palacios-Torres 2017). Although apparently restricted
to the Erythrina genus as host, it
has lots of opportunities. This genus has 116 species distributed across
tropical and subtropical regions: 72 species in the Americas, 31 in Africa, and
12 in Asia.
The
severe damage to wiliwili (and to non-native Erythrina trees planted in urban areas and as windbreaks) prompted Hawaiian
officials to immediately initiate efforts to find a classical biological
control agent. The process moved rapidly. A candidate – a parasitic wasp
species new to science, Eurytoma
erythrinae – was found in East Africa in 2006. Host specificity testing was
carried out. Scientists quickly learned to rear the parasitic wasp in laboratories.
Release of the biocontrol agent was approved in November 2008 – only three and
a half years after the EGW was detected on Oahu.
The
biocontrol agent’s impact was quickly apparent. Establishment was confirmed
within 1–4 months at all release locations throughout Hawai`i. Reduced pest impacts
to trees were detected within two years. By 2018, only 33% of the foliage was
damaged on the majority of wiliwili trees. Damage to non-native Erythrina had also declined.
Results of Biocontrol
Agent’s Release
The
biocontrol agent’s efficacy in reducing EGW’s impacts on trees has been
evaluated for 10 years after the agent’s release. Monitoring was conducted at
sites on four of the six main islands. (The monitoring program and its findings
are described in Kaufman et al. in press).
I wonder how
many other biocontrol agents have been monitored so closely for such a long
time? Shouldn’t they all be?
Given
the uniqueness and importance of such long-term assessment, it is worth looking
at the data in detail.
1) Foliar Damage
and Tree Health
In
2008, before release of the biocontrol agent, more than 70% of young shoots in
wiliwili trees that were inspected were severely infested. The damage rating of
“severe” fell from about 80% of trees in 2008 to about 40% in 2011. About 20%
of trees surveyed – at sites on all islands – had no gall damage.
By
three years after release of the biocontrol agent (2011), mortality rates
attributed to stress from the EGW infestation for trees in natural areas fell
to 21%. Mortality rates for trees in botanical gardens was somewhat higher –
34%. Kaufman et al. proposed several
possible reasons: a) lingering presence of systemic insecticides that might
have harmed the biocontrol agents early in the releases; b) year-round sustenance
for the EGW as a result of the i) presence of alternative hosts and ii) supplemental
irrigation which maintained fresh foliage on the trees.
Less
intensive monitoring occurred during 2013 – 2018. It showed continuing substantial
suppression of EGW damage on Erythrina
foliage, although levels varied among locations. Sites with the lowest
precipitation and higher temperatures throughout the year had the slowest
recovery of wiliwili. Still, trees are now producing vegetative flushes and
healthier canopies during non-dormant periods.
2) Flower and Seed
Damage
Successful
reduction of infestations in flowers and seedpods was less immediate. Still, by
2011, seed-set had increased from less than 3% of trees setting and maturing
seed, to almost 30% with mature seed. The proportion of trees bearing
inflorescences also increased, with more than 60% of trees blooming three years
after introduction of the biocontrol agent. There was also a slow but steady
increase in seed production.
However,
in 2019, it remains unclear how infestation of seedpods will affect germination
and therefore future plant recruitment.
More
worrying, little recruitment was observed over the 10 years. Hawaiian
authorities have completed tests on, and are preparing a petition for release of,
a second biocontrol agent, Aprostocitus
nites. It is hoped that it will further suppress EGW in flowers and
seedpods.
Still, poor recruitment is likely due to the combined impacts of multiple invasive species in native environments. A significant factor is a second insect pest – a bruchid, Specularius impressithorax – which can cause loss of more than 75% of the seed crop. I hope authorities are seeking methods to reduce this insect’s impacts.
The Hawaiian species group of the IUCN has given the wiliwili tree the Reed Book designation of “vulnerable”.
Worries for Naio
naio in bloom photo by Forrest and Kim Starr, courtesy of creative commons
Naio
(Myoporum sandwicense)is an integral component of native Hawaiian
ecosystems, especially in dry forests, lowlands, and upland shrublands.
However, it is also found in mesic and wet forest habitats. Naio is found on all
of the main Hawaiian Islands at elevations ranging from sea level to 3000 m.
The loss of this species would be not only a significant loss of native biological
diversity but also a structural loss to native forest habitats.
The
invasive non-native Myoporum thrips, Klambothrips
myopori, was detected on the Big Island (Hawai‘i Island) in 2009 – four
years after it was first detected on ornamental Myoporum species in California. At the time of the California
detection, the species was unknown to science. It is now known that this
species is native to Tasmania.
The
thrips feeds on and causes galls on plants’ terminal growth and can eventually
lead to death of the plant.
For close to a decade, the Myoporum thrips was restricted to the Big Island. It has now been found on Oahu (Wright pers. comm.) Alarmed by the high mortality of plants in California, in September 2010, the Hawaii Department of Lands and Natural Resources Division of Forestry and Wildlife and the University of Hawai‘i initiated efforts to determine spatial distribution, infestation rates, and overall tree health of naio populations on the Big Island. Monitoring took place at nine protected natural habitats for four years. This monitoring program was supported by the USFS Forest Health Protection program. (See also the chapter on naio by Kaufman et al. 2019 in Potter et al. 2019 – full citation at the end of this blog.)
naio damaged by thrips photo by Leyla Kaufman, University of Hawaii
The monitoring confirmed that the myoporum thrips has spread and colonized natural habitats on the leeward side of Hawai`i Island. Infestation rates increased considerably at all sites over the duration of the four-year sampling period. Trees experiencing high infestation levels also showed branch dieback.
Medium-elevation sites (between 500–999 m) had the highest infestations and dieback: over 70% of the shoots had the worst damage.. At two sites, over 70% of the monitored trees have died.
Even
though flowers and fruits were still seen at all sites, little to no plant
recruitment was observed at these sites. Thus another plant species important
in this endangered plant community is in decline.
Few
management strategies are available for this pest. They include preventing
spread to other islands and early detection followed by rapid application of pesticides.
Implications
and Conclusions
The Erythrina gall wasp and myoporum thrips are only two of the thousands of invasive species established in Hawai`i. Island ecosystems, especially Hawai`i, are well recognized as especially vulnerable to invasive species. It has been estimated that on average 20 new arthropod species become established in Hawai`i every year.
East Africa and Tasmania are new sources for invasive species. Phytosanitary agencies need to adjust their targetting of high-risk imports to recognize this reality. Regarding the Hawaiian introduction of the thrips, there was probably made an intermediary stop in California – which is not unusual. (See also ohia rust.)
I
applaud Hawaiian officials’ quick action to counter these pests. I wish their
counterparts in other states did the same.
There
are multiple threats to Hawaii’s dry forests, including habitat modification
and fragmentation; wild fires; seed predation by rodents; predation on seeds, seedling,
and saplings by introduced ungulates (e.g.
feral goats, pigs and deer); competition with invasive weeds; and damage by
invasive insect pests and diseases.
With
so much of Hawaii’s dry forests already lost, the release of biocontrol agents
targetting specific pests is only one element of a much-needed effort. Long-term protection of wiliwili and naio
depends on greater efforts to reduce all threats and to stimulate natural
regeneration of this ecosystem. These programs could include predator-proof
fencing to keep out ungulates; baiting rodents and snails; and active
collection. Breeding, and planting of threatened plant species in an effort to
protect both the individual species and the habitat.
Kaufman,
L.V., J. Yalemar, M.G. Wright. In press. Classical biological control
of the erythrina gall wasp, Quadrastichus erythrinae, in Hawaii.: Conserving an
endangered habitat. Biological Control. Vol. 142,
March 2020
Potter,
K.M. B.L. Conkling. 2019. Forest Health Monitoring: National Status, Trends,
and Analysis 2018. Forest Service Research & Development Southern Research
Station General Technical Report SRS-239
Kaufman,
L.V, E. Parsons, D. Zarders, C. King, and R. Hauff. 2019. CHAPTER 9. Monitoring
Myoporum thrips, Klambothrips myopori (Thysanoptera: Phlaeothripidae), in
Hawaii
Wright, Mark. 2005. Assistant
Professor and Extension Specialist, University of Hawaii. Personal
communication.
The National Park Service has a legal mandate to manage lands and waters under its jurisdiction so as to “preserve unimpaired” their natural and cultural resources (NPS Organic Act 54 U.S.C. § 100101, et seq.) Invasive species undermine efforts to achieve that mission. In 2000, the NPS adopted a program to coordinate management of invasive plants. It’s not as effective as needed – see the strategic plan.
However, only recently has NPS begun trying to prioritize and coordinate programs targetting the many animals and animal diseases which threaten Park resources. These organisms range from emerald ash borer and quagga mussels; to pythons, goats, and pigs; to diseases such as white nose syndrome of bats and avian malaria in Hawai`i.
In 2017, NPS released an internal study of the pervasive threat to Park resources posed by invasive animals and discussed steps to overcome barriers to more effective responses (Redford et al., 2017; full citation at end of this blog). The Chief of the Biological Resources Division initiated this report by asking a Science Panel to evaluate the extent of the invasive animal problem, assess management needs, review best practices, and assess potential models that could serve as a service-wide organizational framework. The report was to pay particular attention to innovative and creative approaches including, but not limited to, new genomic tools. I summarized the Panel’s findings and conclusions in a blog when its report appeared in 2017.
Significantly, the
Panel’s final report states that “a general record of failure to control
invasive species across the system” was caused principally by a lack of support
for invasive species programs from NPS leadership.
This
report has now appeared in the form of a peer-reviewed article in the journal Biological Invasions by Dayer et al. 2019 (full citation at end of
this blog). Although nine of the ten authors are the same on both reports there
are substantive differences in content. For example, the journal article
reiterates the principal findings and conclusions of the Panel’s final report,
but in less blunt language.
What’s Been Watered
Down
The
toning down is seen clearly in the statements some of the panel’s six key
findings.
Finding
#1
The panel’s report says: invasive animals pose a significant threat to
the cultural and natural values and the infrastructure of U.S. national parks.
To date, the NPS has not effectively addressed the threat they pose.
Dayer et al. says: the ubiquitous presence of invasive animals in parks
undermines the NPS mission.
Finding
#2
The panel’s report says: managing
invasive animals will require action starting at the highest levels, engaging
all levels of NPS management, and will require changes in NPS culture and
capacity.
Dayer et al. says: coordinated action is required to meet the challenge.
Finding
#4
The panel’s report states: effective
management of invasive animals will require stakeholder engagement, education,
and behavior change.
Dayer et al. says: public engagement, cooperation and support is [sic]
critical.
Wording
of the other three “key findings” was also changed, but these changes are less
substantive.
Drayer
et al. also avoid the word “failure”
in describing the current status of NPS” efforts to manage invasive animal
species. Instead, these authors conclude that the invasive species threat “is
of sufficient magnitude and urgency that it would be appropriate for the NPS to
formally declare invasive animals as a service-wide priority.”
Where the
Documents Agree – Sort of
Both
the Panel’s report and Dayer et al.
state that invasive animal threats are under-prioritized and under-funded. They
say that addressing this challenge must begin at the highest levels within the
NPS, engage all levels of management, and will require investments from the NPS
leadership. Even within individual parks,
they acknowledge that staffs struggle to communicate the importance of invasive
animal control efforts to their park leadership, especially given competition with
other concerns that appear to be more urgent. And they admit that parks also
lack staff capacity in both numbers and expertise.
Also,
both the Panel’s report and Dayer et al.
urge the NPS to acknowledge formally that invasive animals represent a crisis
on par with each of the three major crises that drove Service-wide change in
the past: over-abundance of ungulates due to predator control; Yellowstone fire
crisis (which led to new wildfire awareness in the country); and recognition of
the importance of climate change.
The
Panel suggested ways to update NPS’ culture and capacity: providing incentives
for staff to (1) address long-term threats (not just “urgent” ones) and (2) put
time and effort into coordinating with potential partners, including other park
units, agencies at all levels of government, non-governmental organizations,
private landowners, and economic entities. Dayer et al. mention these barriers but does not directly mention
changing incentives as one way to overcome them.
Both
the Panel’s report and Dayer et al.
suggest integrating invasive animal threats and management into long-range
planning goals for natural and cultural landscapes and day-to-day operations of
parks and relevant technical programs (e.g., Biological Resources Division,
Water Resources Division, and Inventory and Monitoring Division).
What is Missing
from the Journal Publication
The
Panel’s final report noted the need for increased funding. It said that such
funding would need to be both consistent and sufficiently flexible to allow
parks to respond to time-sensitive management issues. It proposes several
approaches. These include incorporating some invasive species control programs
(e.g., for weeds and wood borers)
into infrastructure maintenance budgets; adopting invasive species as
fundraising challenges for non-governmental partners (e.g., “Friends of Park” and the National Park Foundation); and
adopting invasive species as a priority threat. Dayer et al. do not discuss funding issues.
The final internal report envisioned the
NPS becoming a leader on the invasive species issue by 1) testing emerging best
management practices, and 2) educating visitors on the serious threat that
invasive species pose to parks’ biodiversity. As part of this process, the
authors suggest that the NPS also take the lead in countering invasive species
denialism. Dayer et al. do not mention the issue of invasive species deniers.
Common Ground:
Status of Invasive Animals in the Parks
The
Panel’s report and Dayer et al. describe
the current situation similarly:
More than half of the National parks that responded to the internal survey (245 of the 326 parks) reported problems associated with one or more invasive animal species.
The total number of species recorded was 331. This is considered to be an underestimate since staffs often lack the ability to thoroughly survey their parks – especially for invertebrates.
Invasive species threats to Parks’ resources have been recognized for nearly 100 years. The original report notes that 155 parks reported the presence of one or more exotic vertebrate species in 1977. At that time, exotic animals were the fourth most commonly reported source of threats. In 1991, parks identified 200 unfunded projects to address exotic species, costing almost $30 million.
Only a small percentage of non-native animal invasions are under active management. Dayer et al. stated that 23% have management plans at the park unit level, and only 11% are reported as being ‘‘under control”.
Individual parks have effective programs targetting specific bioinvaders (examples are described in Redford et al; a brief summary of these efforts is provided in my previous blog.
Common Ground on
Some Solutions
The
report and Dayer et al. promote the
same steps to improve invasive animal management across the Service. Both note
that the NPS is adopting formal decision support tactics to update and
strengthen natural resource management across the board. More specific steps include
establishing
a coordination mechanism that enables ongoing and timely information sharing.
mainstreaming
invasive species issue across the NPS branches or creating a cross-cutting IAS
initiative among the Biological Resources Division, Water Resources Division,
Inventory and Monitoring Division, Climate Change Response Program, and the
regional offices.
While
both documents call on the NPS to develop and test emerging technologies, the
Panel’s final report is more detailed,
providing, in Table 5, a list of several areas of special interest, including
remotely triggered traps, species-specific toxicants, toxicant delivery
systems, drones, environmental DNA, and sterile-male releases. Dayer et al. mention eDNA and metabarcoding
for ED/RR, biocontrol, and gene drives to control invasive pathogens. (Neither
document discusses possible concerns regarding use of CRISPR and other
gene-altering technologies, other than to say there would be public concerns that
would need to be addressed.)
Both
documents note the necessity of working with resource managers beyond park
boundaries to detect and manage species before they arrive in parks. They note
that developing and operationalizing such partnerships requires time and
resources. Furthermore, invasive species prevention, eradication, and
containment programs can be effective only with public support. They suggest
strengthening NPS’ highly regarded public outreach and interpretation program
to build such support, including through the use of citizen scientists.
The
Panel’s final report said that the NPS should recognize that the condition of
the ecosystem is the objective of efforts.
Its authors recognized that achieving this goal might require
reconsidering how ecosystem management is organized within NPS so interacting
stressors (e.g., fire) and management
levers (e.g., pest eradication/suppression, prescribed fire) would be addressed.
For this, the NPS would need to create a focused capacity to address the
pressing issue of invasive animals in such a way that fosters integrated
resource management within parks, focusing on fundamental values of ecosystem
states, and not eradication targets. Dayer et
al. called for the same changes without specifically labelling “condition
of the ecosystem” as the goal.
Publication of
Dayer et al. prompted me to find out
what progress the NPS has made in responding to the “key findings” in the
Panel’s final report (neither publication calls them “recommendations”).
The
National Park Service has acted on the recommendation to appoint an “invasive
animal coordinator” within the Biological Resources Division. That person is Jennifer Sieracki. However, I wonder whether a person located in BRD is of sufficient stature to
influence agency policy across all divisions. It is not clear whether there is active
coordination with the national-level invasive plant coordinator.
Dr.
Sieriaki responded to my query by noting the following new efforts 1) to
improve outreach to partners and
the public, and 2) to expand formal and informal partnerships with local,
state, federal and tribal entities and local communities near parks.
NPS should soon finalize
two formal partnerships with other agencies and organizations for outreach and
management of invasive animal species.
NPS is working with
researchers at the US Geological Survey to expand an existing modeling tool for
identifying potential suitable habitat for invasive plant species to include
invasive insects. This will help staff focus on the most likely locations for
introductions and thus assist with early detection and control.
NPS has created a
Community of Practice so NPS employees can seek each other’s advice on addressing
invasive animal issues. A workshop of regional invasive species coordinators is
planned for the coming months to guide direction of the service-wide program
and identify other top priorities. (Seriacki pers. comm.)
I also wonder whether the NPS can achieve the top-level coordination and outreach to the public called for by both reports while complying with the terms of Public Law 116-9 – the John N. Dingle Jr. Conservation, Management, and Recreation Act, which was enacted a year ago. Title VII, Section 10(i) of this law limits spending to carry out invasive species program management and oversight to 10% of appropriated funds. Less than 15% may be spent on investigations (research), development activities, and outreach and public awareness efforts (Section 10(h)). The law does allow spending for investigations regarding methods for early detection and rapid response, prevention, control, or management; as well as inspections and interception or confiscation of invasive species to prevent in-park introductions.
For more information, see my previous criticism of NPS failure to address invasive species issues here.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
See also my earlier discussion of the new legislation here.
SOURCES
Dayer,
A.A., K.H. Redford, K.J. Campbell, C.R. Dickman, R.S. Epanchin-Niell, E.D.
Grosholz, D.E. Hallac, E.F. Leslie, L.A. Richardson, M.W. Schwartz. 2019. The
unaddressed threat of invasive animals in U.S. National Parks. Biol Invasions
Redford,
K.H., K. Campbell, A. Dayer, C. Dickman, R. Epanchin-Niell, T. Grosholz, D.
Hallac, L. Richardson, M. Schwartz. 2017. Invasive animals in U. S. National Parks:
By a science panel. Natural Resource Report NPS/NRSS/BRD/NRR—2017/1564. NPS,
Fort Collins, Colorado. Commissioned by the NPS Chief of Biological Resources
Division. https://irma.nps.gov/DataStore/DownloadFile/594922
Jennifer Sieracki, Invasive Animal Coordinator,
Biological Resources Division, National Park Service
dead whitebark pine in Crater Lake National Park photo by F.T. Campbell
I began studying and writing about the threat to North America’s forests from non-native insects and pathogens in the early 1990s – nearly 30 years ago. I reported my analyses of the evolving threat in the three “Fading Forests” reports – coauthored by Scott Schlarbaum – in 1994, 2003, and 2014. These reports are available here.
So what has changed over those 30 years? What remains the same? Why have both the changes and the stasis occurred? What can we do to fix the gaps, close unaddressed pathways, strengthen flabby policies? I will address these issues in this and following blogs.
experimental American chestnut planted in Fairfax County, VA photo by F.T. Campbell
What has changed
since the early 1990s:
Adoption and implementation of significant new international and national regulations and programs aimed at preventing introductions of non-native invasive species.
Despite the welter of new regulations, an alarming increase in numbers of highly damaging forest pests established in the country. (By my count, about 50 new species have established on the continent, six on Pacific islands; see details below.)
Alarming spread of established pests to new geographic regions and new hosts (e.g., emerald ash borer in 35 states and 5 provinces; laurel wilt disease across the range of redbay and swamp bay; rapid ‘ōhi‘a death on three of the main Hawaiian islands).
Introductions via unexpected pathways and vectors far removed from phytosanitary agencies’ usual targets, e.g., ship superstructures, imported steel and stone …
What has remained the
same since the early 1990s:
Inadequate
resources provided to response and recovery efforts.
Available
funding focused on only a few of the more than 90 species causing damage.
Adoption
of insufficiently protective regulations that have failed to prevent
introduction and spread of tree-killing pests.
Lengthy
delays in implementing programs that tighten controls – another factor in
continuing introductions and spread.
Continued
importance of expected pathways – nursery stock and raw wood, especially
crates, pallets, and other forms of wood packaging.
Federal
and state agencies still choose not to take action on pests e.g., goldspotted oak borer, polyphagous
and Kuroshio shothole borers, beech leaf disease.
Inadequate
coordination despite several efforts to set priorities.
Spurts
of attention by media and political decision-makers, contrasted by lengthy
periods of inattention.
Failure
of most stakeholders to support efforts to prevent and respond to introductions
of tree-killing pests.
Details: The Situations
Then and Now
(Many of the individual species mentioned here are described more fully here. Full citations of sources are at the end of blog.)
American elm on the National Mall, Washington, D.C.
photo by USDA Agricultural Research Service
In 1993:
The number of non-native forest pest species established in the U.S. was estimated at between 300 (Millers et al. 1993) and 380 (Mattson et al., 1994; Liebhold et al., 1995) .
The area suffering the greatest numbers and impacts was the Northeast.
Several highly damaging pests that had been established for decades, including chestnut blight, white pine blister rust, Port-Orford-cedar root disease, Dutch elm disease, hemlock woolly adelgid, butternut canker, and dogwood anthracnose were receiving some attention but continued to spread.
USDA Forest Service funding for management of exotic pest infestations was crisis-oriented, with “… priorities … set under political pressures for immediate answers, with too much regard for short-term problems and too little consideration for broader management objectives.” (NAS 1975)
Since few high-profile pests had been introduced in recent years, APHIS was not actively engaged. In FY92, APHIS spent $20 million on efforts to eradicate the Asian gypsy moth. The narrow focus is illustrated by the fact that in FY93, more than two-thirds of all USDA tree pest control funds were devoted to efforts to suppress or eradicate the European gypsy moth (See FFI).
Concern about possible new introductions had grown; it focused on proposals to import unprocessed wood from Siberia, New Zealand, and Chile. The USDA Forest Service, academic scientists, and therefore APHIS emphasized the risks of known Asian pests, e.g., Asian gypsy moth, to western coniferous forests (See FFI). While individual scientists had expressed concern about wood packaging material, there was little public discussion of this threat.
We would learn later that several of the most damaging pests were already present in the country but not yet recognized – Asian longhorned beetle, sudden oak death pathogen, probably emerald ash borer.
beech leaf disease
photo by John Pogacnik
In 2019:
Numbers of non-native insects and pathogens attacking trees in North America approach 500 species. (In Fading Forests III, I calculated that by the first decade of the 21st Century, the number had risen to at least 475. Several more have been detected since 2014. More than 181 exotic insects that feed on woody plants had established in Canada. (Source: USDA APHIS. 2000. Wood packaging risk assessment.)
Of these, 91 are considered “serious” threats (Guo et al. 2019). This estimate excludes pests native to portions of North America that are causing severe damage in naïve hosts – e.g., goldspotted oak borer; pests of palms; and pests attacking trees on U.S. Pacific and Caribbean islands.
Introductions had continued.
Between 1980 and 2016, at least 30 non-native species of wood- or bark-boring insects (Scolytinae / Scolytidae) were newly detected in the U.S. (Haack and Rabaglia 2013; Rabaglia et al. 2019). A few of these are highly damaging, e.g. redbay ambrosia beetle, polyphagous and Kuroshio shothole borers.
In addition to these 30 new pests, other highly damaging tree-killing pests probably introduced since the 1980s include (on the continent):
Eight Cerambycids such as Asian longhorned beetle (Wu et al. 2017)
7 Agrilus, including emerald ash borer and soapberry borer; plus goldspotted oak borer transported from Arizona to California (Digirolomo et al. 2019; R. Haack, pers. comm.)
Sirex woodwasp
Pests of palm trees, e.g., red palm mite, red palm weevil, South American palm weevil
Spotted lanternfly
Beech leaf disease
Also not included in the above estimate and lists are tree-killing pests on America’s Pacific Islands :
‘ōhi‘a rust
Cycad scale
Cycad blue betterfly
Erythrina gall wasp
two Ceratocystis pathogens that cause rapid ‘ōhi‘a death
Coconut rhinoceros beetle
Authorities also carried out approximately 25 eradication programs targetting introductions of the Asian gypsy moth (USDA Pest Alert Asian Gypsy Moth plus additional outbreaks since 2014).
Impacts of exacerbated tree mortality rates linked to these introduced pests are seen across wide swaths of the country, and affect widespread species, genera, and families.
dead redbay in Claxton, Georgia photo by Scott Cameron
I will discuss the risk of continuing new introductions in a separate blog.
Trying to Develop
the Big Picture and Set Priorities
In
recent years, USDA Forest Service scientists have made several attempts to
provide nation-wide assessments of the impact of these pests and criteria for
establishing priorities.
The
National Insect and Disease Forest Risk Assessment predicted the loss of basal
area to various pests over the 15-year time period 2012 – 2027. The assessment
predicted the following losses for specific species: 90% for redbay; 60% for whitebark
pine; more than 40% for limber pine; 24% for tanoak; 11% for coast live oak; 6%
for eastern and Carolina hemlock; 27% for eight species of ash; 20% for
American elm; 19% for red oak; 18% for American beech (Krist et al. 2014).
A separate group of scientists found that, nation-wide, non-native forest pests are causing an approximate 5% increase in total mortality by tree volume (Randy Morin at NEFPC). For details on Dr. Morin’s findings, see my blog here.
A third approach to developing a nation-wide picture, Project CAPTURE, (and my blog here) utilized FIA data to develop priorities for conservation action. Fifteen species were placed in the highest priority category, including Florida torreya (Torreya taxifolia), American chestnut and Allegheny and Ozark chinquapins, redbay, five species of ash, two species of hemlock, Port-Orford cedar, tanoak, and butternut (Potter et al. 2019(b).
According
to Project CAPTURE, the non-native pests affecting the largest number of hosts
are the European gypsy moth, which attacks 65 hosts; and oak wilt (Bretziella fagacearum), which infects 61
hosts. The Asian longhorned beetle attacks 43 hosts (Potter et al. 2019(b).
I note that several other non-native pests also have high numbers of host species. In the Project CAPTURE study, these pests are ranked lower because the project limited its evaluation to the five agents with the greatest effect on any particular host. Thus, of the 18 native tree species that host one or both of the invasive shothole borers and associated Fusarium disease complex (PSHB website), the project included only six. Of the 22 tree species listed by APHIS as hosts of Phytophtora ramorum, the project included 12 (K. Potter, pers. comm. April 17, 2019).
SOD-killed tanoak on the Big Sur peninsula, California photo by Matteo Garbelotto, University of California Berkeley
More extensive discussions of
non-native pests’ impacts are provided in Lovett et al. 2006, Lovett et al.
2016, and Potter et al. 2019. A
book-length discussion of invasive species impacts – ranging from feral hogs to
invasive plants, is expected in December; look for Poland et al. (in press).
SOURCES
Aukema,
J.E., D.G. McCullough, B. Von Holle, A.M. Liebhold, K. Britton, & S.J.
Frankel. 2010. Historical Accumulation of Nonindigenous Forest Pests in the
Continental United States. Bioscience. December
2010 / Vol. 60 No. 11
Digirolomo, M.F., E. Jendek, V.V. Grebennikov, O. Nakladal. 2019. First North American
record of an unnamed West Palaearctic Agrilus (Coleoptera:
Buprestidae) infesting European beech (Fagus sylvatica) in New York
City, USA. European Journal of
Entomology. Eur. J.
Entomol. 116: 244-252, 2019
Guo,
Q., S. Fei, K.M. Potter, A.M. Liebhold, and J. Wenf. 2019. Tree diversity
regulates forest pest invasion. Proceedings of the National Academy of Sciences
of the United States of America. www.pnas.org/cgi/doi/10.1073/pnas.1821039116
Haack,
R.A. and R.J. Rabaglia. 2013. Exotic Bark and Ambrosia Beetles in the USA:
Potential and Current Invaders. CAB International 2013. Potential Invasive
Pests of Agricultural Crops (ed. J. Peña)
Krist,
F.J. Jr., J.R. Ellenwood, M.E. Woods, A. J. McMahan, J.P. Cowardin, D.E. Ryerson,
F.J. Sapio, M.O. Zweifler, S.A. Romero 2014. National Insect and Disease Forest
Risk Assessment. United States Department of Agriculture Forest Service Forest
Health Technology Enterprise Team FHTET-14-01
Leung,
B., M.R. Springborn, J.A. Turner, E.G. Brockerhoff. 2014. Pathway-level risk
analysis: the net present value of an invasive species policy in the US. The
Ecological Society of America. Frontiers of Ecology.org
Liebhold, A. M., W. L. MacDonald, D. Bergdahl, and V. C. Mastro. 1995. Invasion by exotic forest pests: a threat to forest ecosystems. Forest Sci., Monograph 30. 49 pp.
Lovett,
G.M., C.D. Canham, M.A. Arthur, K.C. Weathers, and R.D. Fitzhugh. Forest
Ecosystem Responses to Exotic Pests and Pathogens in Eastern North America. BioScience
Vol. 56 No. 5 (May 2006)
Lovett,
G.M., M. Weiss, A.M. Liebhold, T.P. Holmes,
B. Leung, K.F. Lambert, D.A. Orwig, F.T. Campbell, J. Rosenthal, D.G.
McCullough, R. Wildova, M.P. Ayres, C.D. Canham, D.R. Foster, SL. Ladeau, and
T. Weldy. 2016. NIS forest insects and pathogens in the US: Impacts and policy
options. Ecological Applications, 26(5), 2016, pp. 1437–1455
Mattson,
W. J., P. Niemela, I. Millers, and Y. Ingauazo.
1994. Immigrant phytophagous insects on woody plants in the United
States and Canada: an annotated list.
USDA For. Ser. Gen. Tech. Rep. NC-169, 27 pp.
Millers, I. United States
Department of Agriculture, Forest Service Entomologist, Forest Health
Protection Northeastern Area State and Private Forestry. Durham, NH. Personal
communication to F.T. Campbell, 1993.
Morin, R. presentation at Northeastern Forest Pest Council 81st Annual
Meeting, March 12 – 14, 2019, West
Chester, Pennsylvania
National Academy of Sciences. 1975. Forest Pest Control. Washington, D.C.
Poland,
T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019),
Invasive Species in Forests and Grasslands of the United States: A
Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. (in press).
Potter,
K.M., M.E. Escanferla, R.M. Jetton, and G. Man. 2019. Important Insect and
Disease Threats to US Tree Species and Geographic Patterns of Their Potential
Impacts. Forests 2019, 10, 304.
Potter,
K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S. 2019. Prioritizing
the conservation needs of US tree spp: Evaluating vulnerability to forest insect
and disease threats, Global Ecology and Conservation (2019), doi:
https://doi.org/10.1016/
Rabaglia,
R.J., A.I. Cognato, E. R. Hoebeke, C.W. Johnson, J.R. LaBonte, M.E. Carter, and
J.J. Vlach. 2019. Early Detection and Rapid Response. A Ten-Year Summary of the
USDA Forest Service Program of Surveillance for Non-Native Bark and Ambrosia
Beetles. American Entomologist Volume 65, Number 1
U.S.
Department of Agriculture, Animal and Plant Health Inspection Service. 2009.
Risk analysis for the movement of wood packaging material (WPM) from
Canada into the US.
Wu,Y.,
N.F. Trepanowski, J.J. Molongoski, P.F. Reagel, S.W. Lingafelter, H. Nadel1,
S.W. Myers & A.M. Ray. 2017. Identification of wood-boring beetles
(Cerambycidae and Buprestidae) intercepted in trade-associated solid wood
packaging material using DNA barcoding and morphology Scientific Reports 7:40316
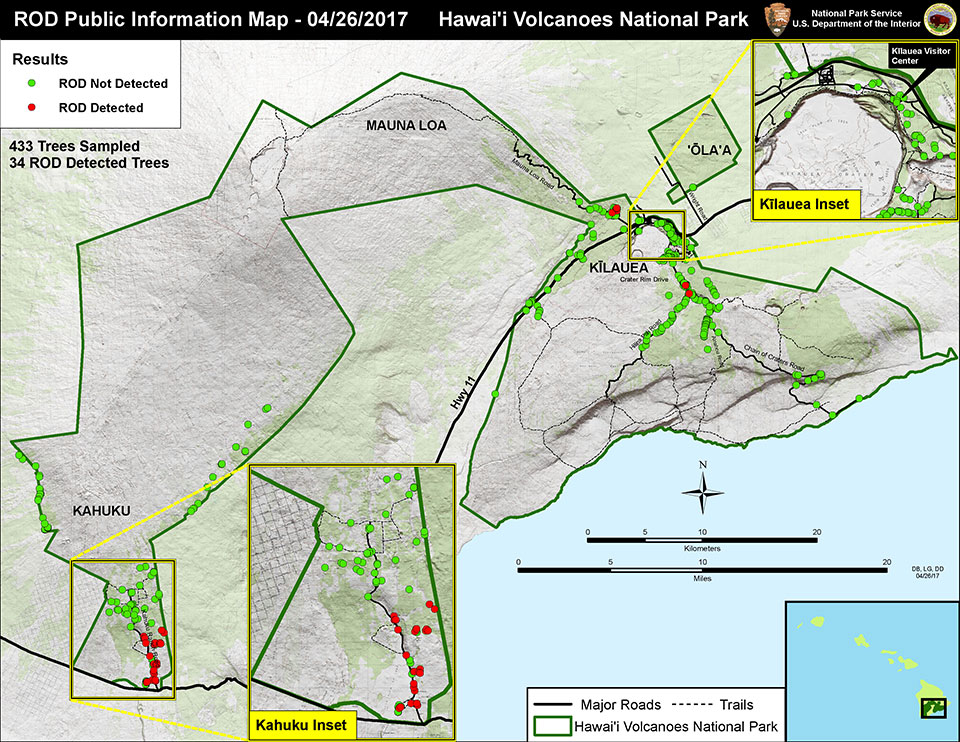



























{kind=link}