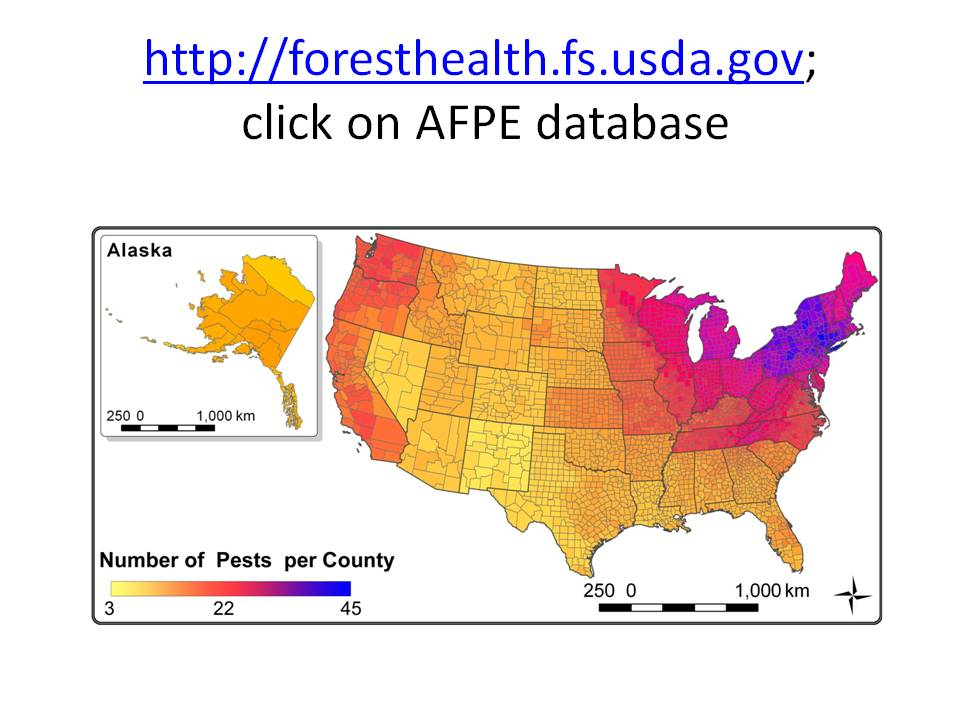
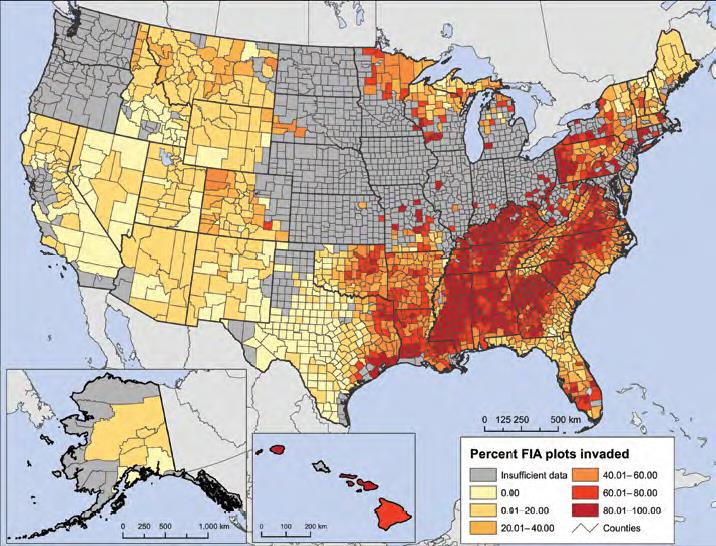
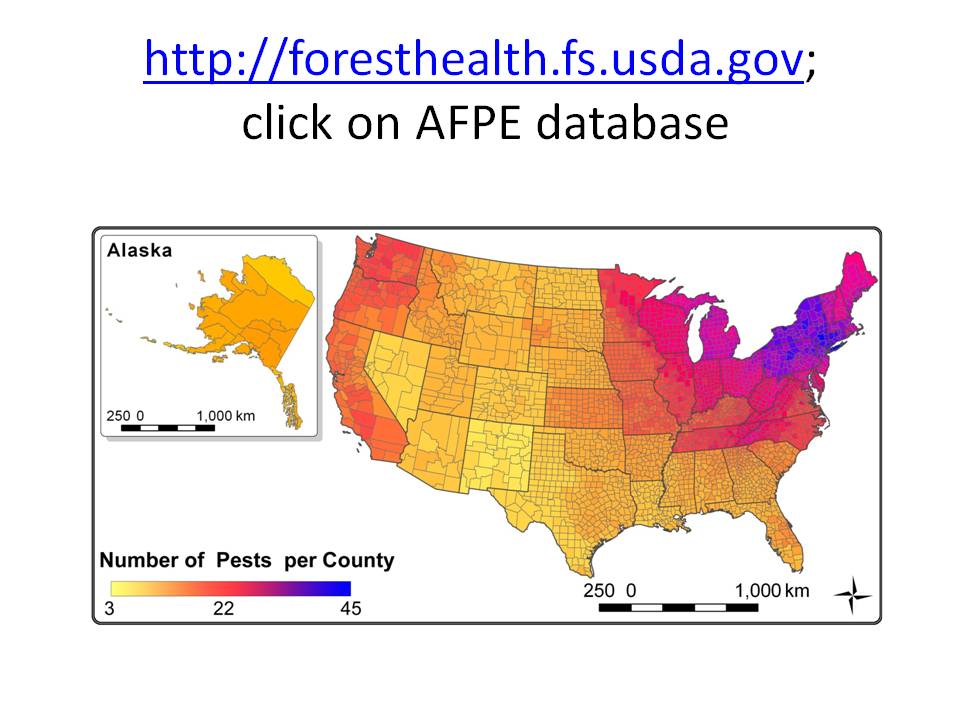
I have posted several blogs recently about tree species’ regeneration. One blog found poor regeneration of many species throughout forests of the eastern United States. Regeneration is particularly poor in the Great Lakes region, western New York and Pennsylvania, along the Mid-Atlantic and New England coasts, and the coastal plain from southern South Carolina to eastern Texas.
A second blog focused on forest succession in New Hampshire. These findings, by Ducey et al., explicitly recognized the impact of non-native tree-killing insects and pathogens. A third article (Payne and Peet, 2023; full citation at the end of this blog) reports similar findings in North Carolina – and explicitly says that the same conditions are found in forests across the eastern United States.
The locations of neither in-depth study – New Hampshire or North Carolina – include those identified by Potter and Riitters (2022) as suffering particularly poor regeneration.
Payne and Peet find that forest succession in the Piedmont region of North Carolina is not proceeding as expected, based on earlier studies conducted in the same region. The differences are apparent at both the canopy and understory levels. Especially notable is the low recruitment of oaks (Quercus species) and hickories (Carya species) – the genera which previous studies indicated would be the climax taxa. One explanation is the disappearance since early in the 20th Century of fire as a driver of disturbance.
The understory communities are also novel, due largely to invasive species: dramatic loss of flowering dogwood (Cornus florida) killed by the non-native pathogen dogwood anthracnose (Discula destructiva), plus overcrowding of the shrub level by invasive plant species. Other drivers are probably suppression of growth of woody species caused by excessive deer herbivory, and overall accelerated shifts in successional trajectory due to hurricane damage.

Forests in eastern North America in the 21st Century face several drivers of change that are either novel or greatly heightened. In addition to the disappearance of chronic fire, these are frequency and timing of hurricanes, feeding by herbivore populations, and introduction of non-native tree-killing pests and plants. Payne and Peet say scientists and managers need to consider these additional drivers – and their interactions! – when anticipating successional change.
Like Ducey et al. in New Hampshire, Payne and Peet used 80 years of data from 33 permanent plots established and 55 years of data from another 3 plots. Twenty-eight of the plots are transitioning from loblolly pine (Pinus taeda) to hardwood dominance; eight plots have been mixed-age hardwood stands since before the study plots were established.
In the North Carolina piedmont, the composition of canopy trees in plots evolving from pine compared to hardwood stands continue to be different 90–120 years after succession began. Canopy trees in upland and bottomland hardwood stands also differ. These differences reflect the relative species in the forest at the initiation of succession dynamics. Hurricanes – especially Hurricane Fran in 1996 – apparently accelerated succession in some plots by toppling the oldest pines. Despite the persistent differences, the species compositions of both canopy and subcanopy layers are trending toward increasing similarity.

The impact of deer browsing is complicated. Deer populations in the study area quadrupled after measurement began in 1980. Deer herbivory suppressed growth of all plant species when their stems were thin (3 – 10 cm DBH). However, after 1996 rapid growth of plants in openings caused by Hurricane Fran’s passage began to reverse the effects of deer browsing. Also, while deer browsing decreases regeneration, growth, and abundance of oak and hickory seedlings and saplings, it also decreases the abundance of other tree species that have – nevertheless – increased in abundance, e.g., red maple (A. rubrum) and black cherry(Prunus serotina).
Payne and Peet found that soil attributes (wetness, texture, organic matter and chemical components), as well as topographic position were minor factors in determining succession trajectories. Increased light availability due to the new or exacerbated drivers of change (thinning of understory vegetation by disease and deer herbivory and opening of the canopy by hurricanes) overcame the influence of nutrients. At most, a unique soil condition might constraining the impacts of these disturbances. Furthermore, these soil-related conditions and other environmental variables change through time — and as a result so does the vegetation. Specifically, the conditions that once supported establishment of oaks and hickories apparently differ today. Payne and Peet conclude that other drivers might be continuing to impact these species’ maturation.
A partial exception is soil nitrogen, through its influence on mycorrhizal patterns. I review mycorrhizal patterns in the discussion of individual tree species, below.
How are Individual Tree Species Responding?
Oaks and hickories are not expanding as expected – either as canopy-sized trees or as seedlings / saplings in the understory. Payne and Peet agree that century-long suppression of low-intensity ground fires is probably the most significant factor in this compositional shift. This decline has been exacerbated by selective logging and deer herbivory. Hickories have established more widely, possibly because young stems have greater shade tolerance. Only plots located on sandy and acidic soils and plots with the greatest hurricane damage have moderate recruitment of oaks and hickories. Oaks and hickories on the poor soils might be aided by the types of ectomycorrhizal fungi that survive in acidic soils with relatively low nitrogen levels. In addition, these soils’ lower water retention probably impedes competition by more mesic, faster-growing, shade-tolerant species. However, even oaks and hickories that have established as seedlings or saplings only rarely progress to canopy dominance. Payne and Peet conclude that oaks might have lost competitive advantage in many of the undisturbed stands.
More mesophytic hardwoods, especially red maple (Acer rubrum), are becoming more numerous and larger – a trend seen throughout forests of the eastern United States. Damage from Hurricane Fran apparently accelerated this trend. However, red maple growth is significantly inhibited by competition from thicket-forming shrubs, especially in bottomland plots. The invasive non-native species thorny olive or oleaster Elaeagnus pungens increased dramatically following Hurricane Fran in 1996. The situation is likely to worsen: two other invasive species, Amur honeysuckle Lonicera maackii and privet Ligustrum japonicum were first detected in the Duke Forest plots in the 2013 survey.
[In New Hampshire, Ducey et al. detected an unexpected levelling off of red maple increases and decline in sugar maple (Acer saccharum); they were unable to determine a cause.]

Another mesophytic hardwood – American beech (Fagus grandifolia) – has become very abundant in bottomland hardwood stands, especially in small-stem size classes in the understory. Beech prefers sandy soils and its ectomycorrhizal associations are apparently more tolerant of more acidic soils.
Payne and Peet mention – briefly and vaguely – uncertainty about the future of beech. The reference cited discusses the impact of beech bark disease (BBD) in the northeast. Range maps indicate that BBD is well established in the southern Appalachians along the North Carolina/Tennessee border; it has apparently not spread as far east as the study area. There is no mention of beech leaf disease (BLD), which is the primary threat to seedlings and saplings. BLD is currently known to be in northern Virginia. It is unknown whether the disease has any climatic or other barrier that would prevent its moving farther south.
Another bottomland indicator taxon that is also increasing in abundance is ash (Fraxinus species). Along with sweetgum (Liquidambar styraciflua), tulip poplar (Liriodendron tulipifera) and black cherry Prunus serotina, ash density and basal area increased dramatically in plots heavily damaged by Hurricane Fran. Payne and Peet expect most ash trees to be killed by emerald ash borer (Agrilus planipennis) by 2022. The beetle was detected in the study area in 2015.

Flowering dogwood (Cornus florida)was one of the most abundant understory species throughout the study area until the late 1980s. The species has declined by more than 80% since then due to the non-native disease dogwood anthracnose (Discula destructiva). No other species has experienced as precipitous a decline. There is now almost no regeneration in most upland sites.
A second species almost eradicated from the study area by a non-native pathogen is American elm (Ulmus americana). Its basal area in 2013 was 5% of peak levels in the 1950s. Most of this loss occurred by the 1960s, shortly after arrived of Dutch elm disease (DED) in North Carolina. A congeneric species, slippery elm U. alata, is reported to beabundant; it is somewhat resistant to DED. There is no mention of the zig-zag sawfly (Aproceros leucopoda) which has been detected in North Carolina, a few counties away from the study area. The foliage-feeding insect’s long-term impact on elm species is not yet understood.
Payne and Peet note that the study area has twice experienced loss of important components due to specialist non-native pathogens: elms and dogwoods. A third similar event looms: ash [The article does not discuss prospects for biological control.] A fourth is less certain: beech. [This numbering assumes that American chestnut and eastern hemlock were not significant components of forests in the study area.] In their view, these events demonstrate the drastic impacts such non-native organisms can have, especially when the host species is highly abundant or otherwise dominant in a specific community. The resulting shifts in community dynamics and modifications to light and water availability due to such losses, can be dramatic and long-lasting, even resulting in novel successional trajectories.

Payne and Peet also emphasize the impact of large, episodic disturbances (in their case, hurricanes). These can have widespread and long-lasting impacts on plant community dynamics. Hurricanes’ frequency, intensity, and timing relative to successional stage are key in determining their impacts on successional trajectories. E.g., strong storms that felled the even-aged pine canopy accelerated succession toward more mixed hardwoods. These changes affect biomass, diversity, competitive dynamics, and invasion by invasive plant species, especially in sites with advantageous soil conditions.
Scientists must also evaluate interactions (both reinforcing and antagonistic) between these drivers. For example, in this study deer herbivory and damage from episodic storms had opposite effects on the density of stems in the understory and therefore the future dynamics of forested stands. Hurricane aftereffects frequently accelerated existing or developing trends resulting from various other drivers (e.g., loss of dogwood to anthracnose disease). [While Ducey et al. also detected lasting impacts from hurricane damage in New Hampshire, these effects did not include changes in tree species composition.] Broader regional and global drivers of change, especially those associated with climate change and nitrogen deposition, interact with these many indicators in novel ways based on their own local loadings.
The Nature Conservancy focuses on fire
The Nature Conservancy magazine for Winter 2023 carries an article describing the organization’s experimental efforts to promote oak succession in the Piedmont forests of North Carolina. Greg Cooper, TNC’s forest ecologist in North Carolina, describes retaining dominance by oaks and hickories – rather than maples and poplars – as vital to protecting the region’s faunal diversity and minimizing impacts from climate change. He says this is because oaks use a quarter of the water of maples and poplars.
Cooper links oaks’ failure to reproduce on fire suppression. TNC kills midstory maples and poplars through hack and squirt methods. This allows more light to penetrate the forest and foster oak seedling recruitment. Then they apply controlled fire. “We currently have 700 acres of [controlled-] burn plots, some of which have been burned twice, some of which have been burned once, [and already] we’re getting more light and an immediate flush of herbaceous diversity. We’re getting a lot more berry species, more wildflowers.” TNC is monitoring plots that have been burned, with and without the pre-burn herbicide treatments, and those that have not been burned. They hope to have results in five to ten years that will indicate whether they are achieving the desired improvement in oak regeneration. If so, they also hope is that in future prescribed burns will be sufficient.
Cooper adds that through the Fire Learning Network and a 23-person fire crew they carry out similar work not just on TNC properties, but also federal and state properties.
SOURCES
Ducey, M.J, O.L., Yamasaki, M. Belair, E.P., Leak, W.B. 2023. Eight decades of compositional change in a managed northern hardwood landscape. Forest Ecosystems 10 (2023) 100121
Payne, C.J. and R.K. Peet. 2023. Revisiting the model system for forest succession: Eighty years of resampling Piedmont forests reveals need for an improved suite of indicators of successional change. Ecological Indicators 154 (2023) 110679
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
or