ash trees killed along Mattawoman Creek in Maryland; photo by Leslie A. Brice
Two important players published documents pressing for “nature-based” solutions to climate change in response to the December 2023 24th Convention of the Parties to the UN Framework Convention on Climate Change.
First, chairs of seven IUCN expert Commissions released a joint statement calling for addressing both climate change and biodiversity loss simultaneously. The elected Commission Chairs represent over 15,000 scientists, scholars, policy makers, economists, lawyers, and other experts who work on issues related to this mission (including me, as well as current and former CISP board members!).
Second, the U.S. Department of the Interior issued detailed guidelines on how to do this.
In this blog, I review the IUCN pronouncement. I will discuss the DOI’s guidelines in a separate blog.
I welcome this statement because I have seen examples of climate “solutions” that worsen the biodiversity crisis. For example, Lugo et al. (2022; full citation at end of this blog) claim to assess the abundance, geographic distribution, contribution to forest structure (including carbon), & temporal trends of non-native tree species. However, they focus almost exclusively on levels of carbon storage. They do not discuss other impacts of non-native tree invasions.
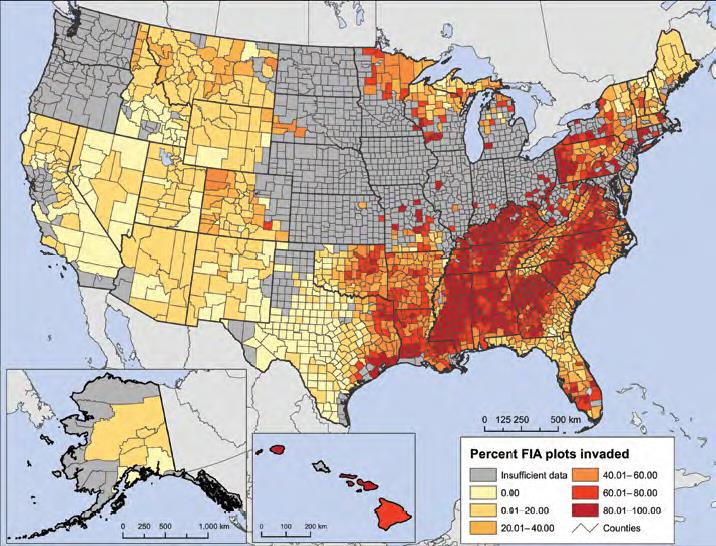
More informative is the 2019 study by Fei et al. ; full citation at end of the blog) that estimated that 41% of total live (woody) biomass in forests of the “lower 48” states was at risk from the most damaging of introduced pests. I pointed out link to blog 159 that elms and beech began dying decades before the underlying (Forest Inventory and Analysis; FIA) data began to be collected. Consequently, the reported mortality rates underestimate the actual loss in biomass associated with these pests. In that blog, I noted that USFS scientists are shifting to new models that will result in a slight bump in overall biomass for the U.S. largely because of increased recognition of the biomass in crowns and limbs. That methodology has now been published.
the “survivor elm” at Longwood Botanical Garden; photo by F.T. Campbell
I also summarized findings by Badgley et al. (2022) that the California cap-and-trade program does not adequately incorporate sequestration losses tied to mortality of tanoak (Notholithocarpus densiflorus) caused by sudden oak death. I noted that California — and North America as a whole – are home to other tree-killing pathogens and insects.
As the IUCN statement clearly demonstrates, climate change and biodiversity loss are inseparable, interdependent, and mutually reinforcing. However, countries’ and businesses’ approaches now fall short of what scientific evidence indicates is needed. We must have bold, transformative, and holistic efforts by scientists – and everyone else.
The IUCN’s full statement has 10 points, which the organization’s blog compresses to four:
1. Integrate Climate and Biodiversity Efforts
The climate and biodiversity challenges require coherent, consistent, and integrated actions that simultaneously limit global warming to a maximum of 1.5oC, conserve and sustainably use biodiversity, and restore degraded ecosystems. Only by considering climate and biodiversity as parts of the same complex, systemic challenge can decision-makers develop effective solutions that maximize benefits while minimizing risks.
“green” infrastructure in urban spaces; Washington, D.C.
2. Enhance Ecosystem Integrity
We humans must maintain, enhance, and restore ecosystem integrity in order to halt biodiversity decline and species extinctions and to maintain the ecosystem services that underpin human well-being. Appropriate actions to conserve and restore terrestrial and marine ecosystems also support climate change mitigation, adaptation, and limits on temperature increases. This is true, however, only as long as chosen actions complement—and are not in lieu of—ambitious reductions of greenhouse gas emissions from fossil fuels, industrial processes, and land-use change.
The full IUCN statement also notes that the effects of “nature-based solutions” must be verified through a robust accounting system. IUCN has released separately a Global Standard for Nature-based Solutions which provides eight specific criteria.
3. Equitably transforming the way we live
Addressing the biodiversity and climate crises will require systemic changes in the way we live. These demand rapid and far-reaching actions across all sectors of a type, scale, and speed never before attempted. IUCN notes, several times, that these transformations must be realized in ways that are equitable and consider impacts on the most vulnerable populations, e.g., indigenous peoples, women, and youth.
IUCN calls for a rapid phase out of fossil fuels, paired with an accelerated and equitable deployment of sustainable clean or renewable energy generation and distribution. In the full statement, IUCN urges countries to avoid relying on unproven — and untested — geoengineering technologies.
4. Prop the Window Open
The window of opportunity to address climate change and biodiversity loss is closing rapidly. Protecting 30% of the Earth’s terrestrial and marine areas by 2030 — a goal adopted by the parties to the Global Biodiversity Convention in late 2022 — will require significant expansion of protected areas in only seven years. I note that while the U.S. is not a party to the biodiversity convention, the Biden Administration has accepted this goal. The IUCN states that achieving this goal depends on greater collaboration across the international agreements on biodiversity, climate change, desertification, and the United Nations’ Sustainable Development Goals. The full statement notes that the United Nations Environment Program (UNEP) calls for tripling expects that funding for nature-based solutions.
old-growth forest in the Pacific Northwest; photo by Richard Orr, via Wikimedia
The IUCN commission chairs warn that delegates at COP28 – and presumably others focused on the climate crisis — must be alert to possible conflicts between biodiversity conservation and climate change mitigation. They cite particularly actions aimed at transitioning energy supplies to “green” sources. This risk arises during choices of sites for solar facilities, wind farms, hydropower dams, and the locations and methods for deep-sea mining for minerals. The IUCN Standard provides guidance for navigating these conflicts.
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. Liebhold. 2019. Biomass losses resulting from insect and disease invasions in US forests. Proceedings of the National Academy of Sciences Vol. 116 No. 35. August 2019
Lugo, A.E., J.E. Smith, K.M. Potter, H. Marcano Vega, and C.M. Kurtz. 2022. The Contribution of NIS Tree Species to the Structure and Composition of Forests in the Conterminous United States in Comparison with Tropical Islands in the Pacific & Caribbean. USDA USFS General Technical Report IITF-54.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
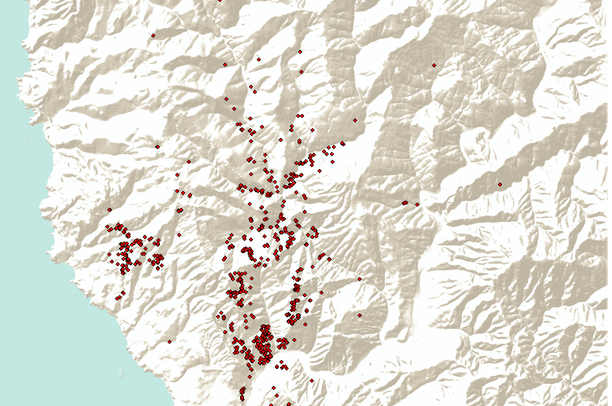
locations of P. ramorum in forests of Oregon in 2023
In a recent blog I offered several critiques of APHIS’ new Phytophthora ramorum risk assessment regarding possible establishment of the causal agent of sudden oak death, in the eastern U.S. states. One of my objections was the brevity of its discussion of the likelihood of sexual combination of the recently introduced EU1 strain with the strain established in North America, NA1 and – more recently – NA2.
This blog provides updates on the status of the Phytophthora ramorum invasion in California and Oregon. My information comes primarily from the newsletter posted by the California Oak Mortality Task Force (COMTF), supplemented by presentations at the recent on-line meeting.
Research by several scientists, including Tyler Bourret, now with USDA Agricultural Research Service, [summarized in the November 2023 COMTF annual meeting] reported that 216 species are now recognized in the genus Phytophthora.
Establishment of Additional Strains of the Pathogen
Scientists now recognize 12 strains of P. ramorum(Sondreli et al., summarized in COMTF newsletter for August 2023). Three of these strains are established in western North American forests. All three – NA1, NA2, & EU1 – are established in southern Curry County, Oregon. Two of the three – EU1 & NA1– are established in neighboring Del Norte County, California. The genetic lineage of the EU1 population in Del Norte points to a link to the Oregon outbreak. [Robinson/Valachovic presentation to COMTF annual meeting November 2023] Given the poor record of efforts to prevent additional introductions of P. ramorum to the United States (the APHIS risk assessment notes that the pathogen has been introduced eight to14 times – or more! — in California), continued introductions of strains not yet established in the U.S. appear likely. Once a strain is established in a North American nursery, it is very likely to spread to nurseries – and possibly forests – in other parts of the country. Remember, the risk assessment reported that P. ramorum has probably been moved over a thousand times on nursery stock from West Coast nurseries across the U.S.
P. ramorum-infected Rhododendron; photo by Jennifer Parke, Oregon State University
Why this matters
Phytophthora ramorum can reproduce sexually only when gametes of the two different mating types (A1 & A2) combine. Most of the North American populations are A2 mating type and most European populations are A1. Establishment of the European EU1 in Oregon and California increases the likelihood that sexual reproduction will occur, which in turn increases the probability that the pathogen will evolve. Sexual combination between NA2 (mating type A2) & EU1 (mating type A1) has occurred at least once – in a nursery in British Columbia. Authorities believe this hybrid has been eradicated. However, the possibility of such matings remains.
The most widespread strain in North America is NA1. It was first detected in the forests north of San Francisco in the middle 1990s; and in Oregon in 2001. Infestations of NA1 are now found from central Curry County, Oregon to Monterey County, California.
The EU1 lineage was first detected in Oregon in 2015. How did it get there since it was previously known only in Europe? The outbreak in Del Norte County, California – detected in 2020 – apparently is associated with the Oregon infection. [Robinson/Valachovic presentation to COMTF annual meeting November 2023] Both states attempted eradication, but the strain is well established. By 2023, the Oregon infestation was detected spreading at sites where intensive surveys in previous years detected no symptomatic trees. In California, new centers of infection have been detected along additional tributary creeks in the area. Scientists expect these infections to spread downhill. Control efforts and even surveys have been hampered by a large fire in the area, which diverted needed personnel and funding. [COMTF newsletter for October 2023 & Robinson/Valachovic]
The NA2 lineage has been found in some nurseries in the Pacific Northwest since 2005. The first detection in forests occurred near Port Orford, Oregon in 2021. Port Orford is 30 miles north of Gold Beach – the hitherto northern extent of the SOD infestation. Oregon authorities believed this signaled a new introduction to the state. By 2023, three sites in the state are now infested with this strain. [Ritokova presentation to COMTF annual meeting November 2023] Oregon now focuses its control efforts on NA2 outbreaks near Port Orford.
In California’s Del Norte County, there are now infestations of two strains of opposite mating types ~ 6 miles apart.The forests between them are conducive to infection, so interactions are likely. Robinson & Valahovic [COMTF annual meeting November 2023] ask how land managers should deal with any interactions. I ask – given the likelihood of hybrids forming – shouldn’t the APHIS risk assessment have tried harder to analyze this risk to the East?
Meanwhile, the NA1 strain continues to spread
In Oregon, the NA1 strain has spread 18 miles to the north and eight miles to the east since 2001 [Ritakova COMTF newsletter October 2023]. In California, spread after the wet winter of 2022-2023 has so far been less than expected. The SOD Blitz [Garbelotto at COMTF annual meeting November 2023] found that the statewide rate of positive trees rose from 7.1% in 2022 to 8.8% in 2023. In the Big Sur region some canyons now test negative that once were positive. Scientists think the negative tests reflect the multi-year drought. Scientists expect the spread will be more visible next year – especially if there is a second wet winter.
As noted above, the exception is in Del Norte County – an area described by CAL FIRE forester Chris Lee as a very wet “pathology” site. SOD (NA1 strain) was first detected in the area north of Crescent City in 2019 [Robinson and Valachovic]. This outbreak could not be re-confirmed for three years, despite intensive surveys. But, in 2022, scientists detected a new concentration of dying tanoak. The infected area is near both rare plants associated with serpentine soils and Jedediah Smith State Park, a unit of Redwood National Park. [Robinson] Meanwhile, the infestation of EU1 strain was first detected in 2020; it has expanded in 2022 and 2023.
In addition to spread facilitated by weather, we also see a continuing role in pathogen transfer via movement of shrubs intended for planting. In fall 2022 Oregon authorities were alerted by a homeowner to an outbreak in Lincoln City, Oregon. This was alarming for four reasons:
it was 201 miles north of the generally infested area in southern in Curry County.
it was well established and had apparently been present for many years.
P. ramorum was not detected in any associated waterways, raising questions about the efficacy of this standard detection method for use in community detections.
one of the infected plants was a new host: western sword fern (Polystichum munitum).
Fortunately, the infection has not (yet) been detected in nearby natural forests. Perhaps this is because there are no tanoaks this far north.
Detection Difficulties
Forest pathologists report several examples of outbreaks involving dozens of trees or plants suddenly being detected in areas which had been surveyed intensively in preceding years with no detections. See Robinson/Valachovic presentation [COMTF annual meeting November 2023, re: both EU1 & NA1 strains in Del Norte County]. I noted above that streams near the Lincoln City, Oregon neighborhood outbreak did not test positive. Nor did water associated with a positive nursery in Oregon[description of Oregon Department of Agriculture nursery regulatory program in COMTF newsletter for August 2023]. Stream baiting is an important component of detection surveys, so I worry about the possible implications of these negative results.
Identification of Additional Hosts [all from COMTF newsletter for August 2023.]
“Mountain Moon” dogwood Cornus capitata [host previously identified in the United Kingdom]
western swordfern (Polystichum munitum) (discussed above)
Oregon P. ramorum eradication attempt; photo by Oregon Department of Forestry
Management
Oregon has tried to manage SOD in the forest since its first detection, but the pathogen’s spread and the recent appearance of two additional strains have overwhelmed the program. One hope was to find a less expensive eradication or containment method. For 20 years, attempts to suppress the disease has focused on eradicating local populations of tanoaks (Notholithocarpus densiflorus) because they are the principal host supporting sporulation in Oregon. When an outbreak has been detected and delimited, they first kill the tanoaks with herbicides to prevent resprouting from the roots. The trees are then felled, piled, and burned. This treatment costs $3,000 – $5,000 / acre. Scientists tested whether they could greatly reduce the cost of the suppression programs by leaving tree boles standing after they have been killed by herbicide. Unfortunately, leaving dead, herbicide-killed trees standing increased sporulation, so this approach would probably exacerbate pathogen spread. [See Jared LeBoldus presentation to COMTF annual meeting November 2023]
Worrying Developments in Europe
In Ireland, sudden larch death – caused by the EU1 strain on Japanese larch (Larix kaempferi) – has spread to several counties. This strain is also causing disease on European beech (Fagus sylvatica) & Noble fir(Abies procera) in locations where these tree grow in association with nearby heavily infected Japanese larch. The EU2 lineage was found in late 2021, infecting L. kaempferi at one site.
Several other Phytophthora species are causing disease on trees, including P. lateralis on Lawson’s cypress, Port-Orford cedar (Chamaecyparis lawsoniana); P. pseudosyringae on Japanese larch; and P. austrocedri on trees in the Juniperus and Cupressus genera.
[information about Ireland from R. O’Hanlon, summarized in COMTF newsletter for August 2023]
Regulation
The European Union has relaxed phytosanitary regulation of Phytophthora ramorum. Previously the species – all strains – was considered a quarantine pest. Now its regulatory status depends on the origin of the infected material. “Non-EU isolates” of Phytopththora ramorum are still quarantine pests (presumably the two North American strains [NA1 & NA2] and the eight other strains identified in Asian forests). These pests are treated as the most serious pests in the Union; when they are detected, extensive control actions must be taken. “EU isolates” (presumably EU1 & EU2) are now treated as regulated non-quarantine pests. The focus is to limit the spread of these on plants for planting only.
The European Union and USDA APHIS regulatory emphases differ to some extent (APHIS does not regulate P. ramorum in natural settings, only interstate movement via, inter alia, the nursery trade). However, I am worried that both seem intent on minimizing their regulatory programs.
Arbutus canariensis; photo by Moreno José Antonio via Plantnet
Another region at risk
Macaronesia is a group of several North Atlantic islands,e.g., Madeira and the Azores, Canary, and Cape Verde islands.The islands have climates similar to areas affected by P. ramorum. The Macaronesian laurel forest is a remnant subtropical evergreen forest which shares some plant taxa with those that host the pathogen elsewhere. Moralejo et al. found that, overall, plant species showed considerable tolerance of the pathogen. However, P. ramorum was “rather aggressive” on Viburnum tinus, Arbutus canariensis and Ilex canariensis. Furthermore, mean sporangia production on five Macaronesian laurel forest species was similar to levels on Umbellularia californica, a key host driving the SOD epidemics in California.Moralejo et al. concluded that there is a moderate to high risk of establishment if Phytophthora ramorum were introduced in the Macaronesian laurel forest. [Study summarized in October 2023 COMTF newsletter.]
Important Research
The COMTF August newsletter reports exciting work developing improved detection tools for Phytophthora species, especially P. ramorum. Sondreli, Tabima, & LeBoldus have developed a method to quickly distinguish among the four most common clonal lineages (NA1, NA2, EU1 and EU2). These assays are sensitive to weak concentrations and effective in testing a variety of sample types including plant tissue and cultures. Oregon State University is already using in its diagnostic laboratory.
YuFang, Xia, Dai, Liu, Shamoun, and Wu have developed a simple, rapid, sensitive detection system for the molecular identification of P. ramorum that does not require technical expertise or expensive ancillary equipment. It can be used in laboratory or using samples collected from the field.
Quiroga et al. found that thinning – with or without burning of the slash – significantly reduced stand density and increased average tree size without significantly decreasing total basal area. This effect persisted for five years after treatments – especially when supported by follow-up basal sprout removal. Preventative treatments also significantly increased dominance of tree species not susceptible to Phytophthora ramorum.
In a study summarized in the October 2023 COMTF newsletter, Bourret et al. reported results of nearly 20 years of leaf baiting in watersheds covering an 800-mile section of the Pacific Coast in northern and central California. They found 22 Phytophthora & Nothophytophthora species.Several – including P. ramorum — were abundant and widespread. Some isolates in northern California differ from those found elsewhere. Mitochondrial sequences revealed multiple hybridization events between P. lacustris and P. riparia.
Bourret et al. also found that P. pluvialis is probably native to Western North America. The strain invasive on conifers in New Zealand probably originated in California rather than Oregon or Washington.
Jared LeBoldus and colleagues are studying the ecological impact of tanoak mortality in Oregon forests. [Summarized in November 2023 COMTF newsletter.] They expect impacts at various trophic levels and functions. Preliminary findings regarding the plant community show increases in understory and herbacious species diversity; a shift away from tanoak to Douglas-fir; and increased coarse woody debris. These findings are similar to results from studies in central California by Dave Rizzo and colleagues at UC Davis. LeBoldus is now studying the microbiome of plant leaves; soil mycorrhizal diversity; invertebrates and pollinators (loss of the large annual flower crop of tanoaks presumably affects pollinators). They hope in the future to study small mammal communities (which they expect to be affected by the loss of acorns).
Jared LeBoldus and colleagues also reported early results of genomic studies exploring disease resistance in tanoaks. Various scientists started such studies in the past, but so far all efforts have petered out due to absence of sustained funding, support from agency management, and links to facilities with the necessary tree improvement/breeding resources. (See Richard Sneizko’s description of requirements for resistance breeding, here.) I hope this project proves more sustainable.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
hardwood regeneration in northern Virginia forest; photo F.T. Campbell
I have posted several blogs recently about tree species’ regeneration. One blog found poor regeneration of many species throughout forests of the eastern United States. Regeneration is particularly poor in the Great Lakes region, western New York and Pennsylvania, along the Mid-Atlantic and New England coasts, and the coastal plain from southern South Carolina to eastern Texas.
A second blog focused on forest succession in New Hampshire. These findings, by Ducey et al., explicitly recognized the impact of non-native tree-killing insects and pathogens. A third article (Payne and Peet, 2023; full citation at the end of this blog) reports similar findings in North Carolina – and explicitly says that the same conditions are found in forests across the eastern United States.
The locations of neither in-depth study – New Hampshire or North Carolina – include those identified by Potter and Riitters (2022) as suffering particularly poor regeneration.
Payne and Peet find that forest succession in the Piedmont region of North Carolina is not proceeding as expected, based on earlier studies conducted in the same region. The differences are apparent at both the canopy and understory levels. Especially notable is the low recruitment of oaks (Quercus species) and hickories (Carya species) – the genera which previous studies indicated would be the climax taxa. One explanation is the disappearance since early in the 20th Century of fire as a driver of disturbance.
The understory communities are also novel, due largely to invasive species: dramatic loss of flowering dogwood (Cornus florida) killed by the non-native pathogen dogwood anthracnose (Discula destructiva), plus overcrowding of the shrub level by invasive plant species. Other drivers are probably suppression of growth of woody species caused by excessive deer herbivory, and overall accelerated shifts in successional trajectory due to hurricane damage.
flowering dogwood autumn display; F.T. Campbell
Forests in eastern North America in the 21st Century face several drivers of change that are either novel or greatly heightened. In addition to the disappearance of chronic fire, these are frequency and timing of hurricanes, feeding by herbivore populations, and introduction of non-native tree-killing pests and plants. Payne and Peet say scientists and managers need to consider these additional drivers – and their interactions! – when anticipating successional change.
Like Ducey et al. in New Hampshire, Payne and Peet used 80 years of data from 33 permanent plots established and 55 years of data from another 3 plots. Twenty-eight of the plots are transitioning from loblolly pine (Pinus taeda) to hardwood dominance; eight plots have been mixed-age hardwood stands since before the study plots were established.
In the North Carolina piedmont, the composition of canopy trees in plots evolving from pine compared to hardwood stands continue to be different 90–120 years after succession began. Canopy trees in upland and bottomland hardwood stands also differ. These differences reflect the relative species in the forest at the initiation of succession dynamics. Hurricanes – especially Hurricane Fran in 1996 – apparently accelerated succession in some plots by toppling the oldest pines. Despite the persistent differences, the species compositions of both canopy and subcanopy layers are trending toward increasing similarity.
deer-damaged red maple; photo by Eli Sagor via Flickr
The impact of deer browsing is complicated. Deer populations in the study area quadrupled after measurement began in 1980. Deer herbivory suppressed growth of all plant species when their stems were thin (3 – 10 cm DBH). However, after 1996 rapid growth of plants in openings caused by Hurricane Fran’s passage began to reverse the effects of deer browsing. Also, while deer browsing decreases regeneration, growth, and abundance of oak and hickory seedlings and saplings, it also decreases the abundance of other tree species that have – nevertheless – increased in abundance, e.g., red maple (A. rubrum) and black cherry(Prunus serotina).
Payne and Peet found that soil attributes (wetness, texture, organic matter and chemical components), as well as topographic position were minor factors in determining succession trajectories. Increased light availabilitydue to the new or exacerbated drivers of change (thinning of understory vegetation by disease and deer herbivory and opening of the canopy by hurricanes) overcame the influence of nutrients. At most, aunique soil condition might constraining the impacts of these disturbances. Furthermore, these soil-related conditions and other environmental variables change through time — and as a result so does the vegetation. Specifically, the conditions that once supported establishment of oaks and hickories apparently differ today. Payne and Peet conclude that other drivers might be continuing to impact these species’ maturation.
A partial exception is soil nitrogen, through its influence on mycorrhizal patterns. I review mycorrhizal patterns in the discussion of individual tree species, below.
How are Individual Tree Species Responding?
Oaks and hickories are not expanding as expected – either as canopy-sized trees or as seedlings / saplings in the understory. Payne and Peet agree that century-long suppression of low-intensity ground fires is probably the most significant factor in this compositional shift. This decline has been exacerbated by selective logging and deer herbivory. Hickories have established more widely, possibly because young stems have greater shade tolerance. Only plots located on sandy and acidic soils and plots with the greatest hurricane damage have moderate recruitment of oaks and hickories. Oaks and hickories on the poor soils might be aided by the types of ectomycorrhizal fungi that survive in acidic soils with relatively low nitrogen levels. In addition, these soils’ lower water retention probably impedes competition by more mesic, faster-growing, shade-tolerant species. However, even oaks and hickories that have established as seedlings or saplings only rarely progress to canopy dominance. Payne and Peet conclude that oaks might have lost competitive advantage in many of the undisturbed stands.
More mesophytic hardwoods, especially red maple(Acer rubrum), are becoming more numerous and larger – a trend seen throughout forests of the eastern United States. Damage from Hurricane Fran apparently accelerated this trend. However, red maple growth is significantly inhibited by competition from thicket-forming shrubs, especially in bottomland plots. The invasive non-native species thorny olive or oleasterElaeagnus pungensincreased dramatically following Hurricane Fran in 1996. The situation is likely to worsen: two other invasive species, Amur honeysuckle Lonicera maackii and privet Ligustrum japonicum were first detected in the Duke Forest plots in the 2013 survey.
[In New Hampshire, Ducey et al. detected an unexpected levelling off of red maple increases and decline in sugar maple (Acer saccharum); they were unable to determine a cause.]
beech-dominated understory in northern Virginia; F.T. Campbell
Another mesophytic hardwood – American beech(Fagus grandifolia) – has become very abundant in bottomland hardwood stands, especially in small-stem size classes in the understory. Beech prefers sandy soils and its ectomycorrhizal associations are apparently more tolerant of more acidic soils.
Payne and Peet mention – briefly and vaguely – uncertainty about the future of beech. The reference cited discusses the impact of beech bark disease (BBD) in the northeast. Range maps indicate that BBD is well established in the southern Appalachians along the North Carolina/Tennessee border; it has apparently not spread as far east as the study area. There is no mention of beech leaf disease (BLD), which is the primary threat to seedlings and saplings. BLD is currently known to be in northern Virginia. It is unknown whether the disease has any climatic or other barrier that would prevent its moving farther south.
Another bottomland indicator taxon that is also increasing in abundance is ash (Fraxinus species). Along with sweetgum (Liquidambar styraciflua), tulip poplar (Liriodendron tulipifera) and black cherry Prunus serotina, ash density and basal area increased dramatically in plots heavily damaged by Hurricane Fran. Payne and Peet expect most ash trees to be killed by emerald ash borer (Agrilus planipennis) by 2022. The beetle was detected in the study area in 2015.
ash killed by EAB on Potomac lowlands; F.T. Campbell
Flowering dogwood (Cornus florida)was one of the most abundant understory species throughout the study area until the late 1980s. The species has declined by more than 80% since then due to the non-native disease dogwood anthracnose (Discula destructiva). No other species has experienced as precipitous a decline. There is now almost no regeneration in most upland sites.
A second species almost eradicated from the study area by a non-native pathogen is American elm (Ulmus americana). Its basal area in 2013 was 5% of peak levels in the 1950s. Most of this loss occurred by the 1960s, shortly after arrived of Dutch elm disease (DED) in North Carolina. A congeneric species, slippery elm U. alata, is reported to beabundant; it is somewhat resistant to DED. There is no mention of the zig-zag sawfly (Aproceros leucopoda) which has been detected in North Carolina, a few counties away from the study area. The foliage-feeding insect’s long-term impact on elm species is not yet understood.
Payne and Peet note that the study area has twice experienced loss of important components due to specialist non-native pathogens: elms and dogwoods. A third similar event looms: ash [The article does not discuss prospects for biological control.] A fourth is less certain: beech. [This numbering assumes that American chestnut and eastern hemlock were not significant components of forests in the study area.] In their view, these events demonstrate the drastic impacts such non-native organisms can have, especially when the host species is highly abundant or otherwise dominant in a specific community. The resulting shifts in community dynamics and modifications to light and water availability due to such losses, can be dramatic and long-lasting, even resulting in novel successional trajectories.
Members of the 23rd Civil Engineer Squadron/23rd Wing chainsaw a tree lying across a street in the NCO housing area- damage to piedmont North Carolina by Hurricane Fran. Photo courtesy of U.S. National Archives.
Payne and Peet also emphasize the impact of large, episodic disturbances (in their case, hurricanes). These can have widespread and long-lasting impacts on plant community dynamics. Hurricanes’ frequency, intensity, and timing relative to successional stage are key in determining their impacts on successional trajectories. E.g., strong storms that felled the even-aged pine canopy accelerated succession toward more mixed hardwoods. These changes affect biomass, diversity, competitive dynamics, and invasion by invasive plant species, especially in sites with advantageous soil conditions.
Scientists must also evaluate interactions (both reinforcing and antagonistic) between these drivers. For example, in this study deer herbivory and damage from episodic storms had opposite effects on the density of stems in the understory and therefore the future dynamics of forested stands. Hurricane aftereffects frequently accelerated existing or developing trends resulting from various other drivers (e.g., loss of dogwood to anthracnose disease). [While Ducey et al. also detected lasting impacts from hurricane damage in New Hampshire, these effects did not include changes in tree species composition.] Broader regional and global drivers of change, especially those associated with climate change and nitrogen deposition, interact with these many indicators in novel ways based on their own local loadings.
The Nature Conservancy focuses on fire
The Nature Conservancy magazine for Winter 2023 carries an article describing the organization’s experimental efforts to promote oak succession in the Piedmont forests of North Carolina. Greg Cooper, TNC’s forest ecologist in North Carolina, describes retaining dominance by oaks and hickories – rather than maples and poplars – as vital to protecting the region’s faunal diversity and minimizing impacts from climate change. He says this is because oaks use a quarter of the water of maples and poplars.
Cooper links oaks’ failure to reproduce on fire suppression. TNC kills midstory maples and poplars through hack and squirt methods. This allows more light to penetrate the forest and foster oak seedling recruitment. Then they apply controlled fire. “We currently have 700 acres of [controlled-] burn plots, some of which have been burned twice, some of which have been burned once, [and already] we’re getting more light and an immediate flush of herbaceous diversity. We’re getting a lot more berry species, more wildflowers.” TNC is monitoring plots that have been burned, with and without the pre-burn herbicide treatments, and those that have not been burned. They hope to have results in five to ten years that will indicate whether they are achieving the desired improvement in oak regeneration. If so, they also hope is that in future prescribed burns will be sufficient.
Cooper adds that through the Fire Learning Network and a 23-person fire crew they carry out similar work not just on TNC properties, but also federal and state properties.
SOURCES
Ducey, M.J, O.L., Yamasaki, M. Belair, E.P., Leak, W.B. 2023. Eight decades of compositional change in a managed northern hardwood landscape. Forest Ecosystems 10 (2023) 100121
Payne, C.J. and R.K. Peet. 2023. Revisiting the model system for forest succession: Eighty years of resampling Piedmont forests reveals need for an improved suite of indicators of successional change. Ecological Indicators 154 (2023) 110679
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
affects of mountan pine beetle on lodgepole pine in Rocky Mountain National Park, Colorado photo from Wikimedia; one of pests addressed by USFS FHP
Several USFS scientists have published an assessment of the agency’s program to enhance forest health across the country: the Forest Health (FHP) program. [see Coleman et al., full citation at end of this blog.] The program assists cooperators (including other federal agencies) to prevent, suppress, and eradicate insect and pathogen outbreaks affecting trees, regardless of land ownership.
Each year, I advocate for adequate funding for the FHP program — which comes from annual Congressional appropriations. Funding has remained static at about $100 million per year. I interpret the article as providing support for my call for increased appropriations. First, it reports that the number of projects and extent of area treated have declined from 2011 to 2020. This is because static funding levels are stretched increasingly thin as costs to implement the same activities rise. Second, the program does not address many damaging forest pests already in the country. The result is growth of established threats to forest health. Finally, new insects and pathogens continue to be introduced. Protecting forest health necessitates tackling these new pests – and that requires money and staff.
Coleman et al. analyzed data from the decade 2011- 2020 to determine the most frequently used project types, integrated pest management (IPM) strategies and tactics, dominant forest pests and associated hosts managed, and most comprehensive forest IPM programs in practice. While there is a wide range of possible projects, most of those funded consist of some form of treatment (more below). The databases relied on do not include funding through the National Forest System aimed at improving forest health through such management activities as stand thinning treatments and prescribed fire. Nor are all pest management activities recorded in the centralized databases. I regret especially the fact that “genetic control” (= resistance breeding) are left out.
Port-Orford cedar seedlings in trial for resistance to Phytophthora lateralis at Dorena center; photo courtesy of Richard Sniezko, USFS
Summary of Findings
The data are sorted in various categories, depending on whether one wishes to focus on the type of organism being managed or the management approach. All presentations make evident a dramatic imbalance in the projects funded. Again and again, spongy moth (Lymantria dispar dispar), southern pine beetle (SPB, Dendroctonus frontalis), and several bark beetles attacking conifers in the West (in particular mountain pine beetle, [MPB] Dendroctonus ponderosae) dominate, as measured by both funding and area treated.
oak trees in Shenandoah National Park killed by spongy moth; photo by F.T. Campbell
The bulk of the funding went to the above species, plus hemlock woolly adelgid (HWA; Adelges tsugae); emerald ash borer (EAB, Agrilus planipennis), oak wilt (caused by Bretziella fagacearum), and white pine blister rust (WPBR, Cronartium ribicola).
95% of the projects focused on only four taxa: oaks, Quercus spp. [spongy moth suppression and eradication]; loblolly and ponderosa pines [bark beetle prevention and suppression]; and eastern hemlock [HWA suppression].
Projects seeking to suppress an existing pest outbreak covered 87% of the total treatment area. However, 98% of the treated area was linked to only 20 taxa; again, spongy moth dominated.
Projects seeking to prevent introduction or spread of a pest constituted only 30% of all projects and covered only 11% of the total treatment area.
Eradication and restoration projects each equaled less than 5% of total projects and treatment areas.
Native forest pests were targetted by 79% of projects; non-native pests by 21%. However, non-native pests accounted for 84% of the total treatment area (again, the spongy moth).
While 67% of projects took place on USFS lands (focused on MPB and SPB), 89% of the total treatment area was on lands managed by others (state or other federal agencies, or private landowners). Again, the size of the non-USFS area treated was driven primarily by the spongy moth Slow the Spread program.
Insect pests received nearly all of the funding: 70% of funding targetted phloem-feeding insects, especially SPB and MPB; 10% targetted foliage feeders, especially spongy moth; 6% targetted sap feeders. 4% tackled rusts (e.g., WPBR); just 2% addressed wood borers (e.g., Asian longhorned beetle, emerald ash borer).
The ranking by size of area treated differs. In this case, 82% of areas treated face damage by foliage feeders (e.g., spongy moth); 15% of the treated areas are threatened by phloem feeders (e.g., MPB); only 1.4% of the area is damaged by sap feeders (e.g., HWA); 0.6% is threatened by rust; and 0.2% by wood borers.
Re: control strategies, 32% of projects relied on silvicultural strategies; 22% used semiochemical strategies; 21% exploited other chemical controls; and 18% used physical/mechanical control methods.
Coleman et al. regretted that few programs incorporated microbial/biopesticide control strategies; these were applied on only 10% of total treated area. Again, the vast majority of such projects were aerial applications of spongy moth controls, Bacillus thuringiensis var. kurstaki (Btk) and nucleopolyhedrosis viruses (NPV) (Gypchek). Coleman et al. called for more research to support this approach efforts to overcome other obstacles (see below).
Coleman et al. also called for better record-keeping to enable analysis of genetic control/ resistance breeding projects, treatment efficacy, and survey and technical assistance activities.
History
The article provides a brief summary of the history of the Forest Service’ pest management efforts. Before the 1960s, the USFS relied on labor-intensive physical control tactics, classical biocontrol, and widespread chemical applications. Examples include application of pesticides to suppress or eradicate spongy moth; decades of Ribes removal to curtail spread of white pine blister rust; salvage logging and chemical controls to counter phloem feeders / bark beetles in the South and West. These strategies were increasingly replaced by pest-specific management tactics during the 1970s.
Over the decade studied (2011-2020), tree defoliation attributed to various pests (including pathogens) affected an estimated 0.7% of the 333 million ha of U.S. forest land annually. Mortality attributed to pests impacted an estimated 0.8% of that forest annually. See Table 1. Two-thirds of the area affected by tree mortality is attributed to phloem feeders; a distant second agent is wood borers. These data are incomplete because many insects, diseases, and parasitic higher plants are not tracked by aerial surveys.
As I noted above, these data do not include projects that screen tree species to identify and evaluate genetic resistance to a pest; or efforts to collect cones, seed, and scion. I consider these gene conservation and resistance programs to be some of the most important pest-response efforts. I have blogged about the USFS’ Dorena Genetic Resource Center’ efforts to breed five-needle pines, Port-Orford cedar, and ash. link
41% of silvicultural control treatments targetted phloem feeders; 48% addressed cankers and rusts together. Restoration planting was done in response to invasions by ALB, EAB, and WPBR, as well as native bark beetles and mistletoes.
effort to eradicate SOD in southern Oregon; partially funded by USFS FHP. Photo courtesy of Oregon Department of Forestry
Physical/mechanical control projects were most widely applied in the Rocky Mountains in response particularly to diseases: vascular wilts, rusts, and cankers, including WPBR. This type of project was also used to deal with non-native diseases in other parts of the country, e.g., oak wilt, sudden oak death (SOD), Port-Orford cedar root rot, and rapid ʻōhiʻa death. Sanitation treatments (i.e., removal of infected/infested trees) was used for native mistletoes and root rots, and some non-native insects, e.g., EAB and coconut rhinoceros beetle (Oryctes rhinoceros). Pruning is a control strategy for WPBR. Trenching is applied solely to suppress oak wilt.
Chemical controls were limited to small areas. These projects targetted seed/cone/flower fruit feeders, foliage and shoot diseases, sap feeders [e.g., balsam woolly adelgid (BWA), HWA], wood borers (e.g., EAB) and phloem feeders (e.g., Dutch elm disease; DMF oak wilt vectors). Cover sprays have been used against goldspotted oak borer (GSOB); and many native insects. Fungicides are rarely used; some is applied against the oak wilt pathogen in areas inaccessible by heavy equipment.
treating hemlock trees in Conestee Falls, NC; photo courtesy of North Carolina Hemlock Restoration Initiative
Classical biocontrol projects funded by the program targetted almost exclusively HWA. Some 4.3 million predators have been released since the early 1990s; 820,057 in just the past 10 years.
Gene conservation and breeding projects were directed primary at commercially important hosts, e.g., loblolly Pinus taeda and slash pine P. elliottii; and several non-native pests, including chestnut blight, EAB, HWA, and WPBR.
Survey and technical assistance (i.e., indirectly funded activities) conducted by federal, state, and tribal personnel contributed to education/outreach, evaluating effectiveness, identification, monitoring, and record keeping strategies.
As should be evident from the data presented here, suppression treatments dominated by number of projects and treatment area. The poster child project is the national spongy moth Slow the Spread program. The authors say this program is the most advanced forest IPM program in the world. It has successfully slowed spongy moth’s rate of spread by more than 80% for more than 20 years.
A second widely-used subset of suppression programs consists of physical / mechanical control. This is often the principal suppression strategy in high-visitation sites (e.g., administration sites, campgrounds, picnic areas, and recreation areas). Sanitation harvests are one of the few viable management techniques for suppressing or slowing the spread of recently introduced non-native pests. Nevertheless, the largest number of suppression projects and use of sanitation treatments focused on a native pest, mountain pine beetle, at the height of its outbreak in early 2010s.
Silvicultural control, specifically tree thinning, represents the predominant forest pest prevention tactic, especially on lands managed by the USFS. Two programs dominate: the Southern Pine Beetle Prevention Program and the Western Bark Beetle Initiative. Again, Coleman et al. assess these treatments as very successful. Forest thinning treatments also address other management concerns, i.e., reduce threat of catastrophic wildfires and reduce adverse effects of climate change.
Chemical control tactics are applied to suppress most forest insect feeding guilds in high-value sites and seed orchards. Soil or tree injections of systemic pesticides are used to protect ash and hemlock trees. Topical sprays have been applied to protect whitebark pine (Pinus albicaulis) from mountain pine beetle. Whitebark pine was listed as threatened under the Endangered Species Act in December 2022.
dead whitebark pine at Crater Lake NP; photo by F.T. Campbell
Soil or tree injections target two non-native insects, EAB and HWA.
Genetic control via resistance breeding represents the primary strategy to combat several non-native diseases. (More options are typically available for insects than diseases.) Coleman et al. focus on the extensive effort to protect many of the five-needle pines from WPBR. As I have described in earlier blogs, the Dorena Genetic Resource Center in Oregon has engaged on numerous other species, too.
Coleman et al. describe pest-management associated monitoring efforts as consisting largely of coordinated annual aerial detection surveys, detection trapping, stream-baiting ofPhytophthora ramorum, and ground surveys to address site-specific issues.
Coleman et al. call for improvement of record-keeping / databases to encompass all pests, management actions, and ownerships. They also advocate for additional decision-making tools, development of microbial/biopesticides, genetic research and breeding, and biocontrol strategies for several pest groups.
They consider the southern pine beetle and spongy moth programs to be models of comprehensive IPM programs that could be adapted to additional forest health threats. They note, however, that development and implementation of these programs require significant time, financial commitments, and collaborations from various supporting agencies. Not all programs enjoy such resources.
SOURCE
Coleman, T.W, A.D. Graves, B.W. Oblinger, R.W. Flowers, J.J. Jacobs, B.D. Moltzan, S.S. Stephens, R.J. Rabaglia. 2023. Evaluating a decade (2011–2020) of integrated forest pest management in the United States
Journal of Integrated Pest Management, (2023) 14(1): 23; 1–17
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Rome: home of the International Plant Protection Convention
I often assert that the international phytosanitary system has proven to be a failure in preventing introductions.
Some of the recent publications support my conclusion – although most don’t say so explicitly. For example, the Fenn-Moltu et al. (2023) study of insect transport and establishment around the world found that the number of invasive species-related treaties, regulations and legislation a country has adopted had no significant effect on either the number of insect species detected at that country’s border or the number of insect species that established in that country’s ecosystems..
Weber et al. also found considerable evidence that international and U.S. phytosanitary systems are not curtailing introduction of insects and entomophagic pathogens. In my earlier blog I review their study of unintentional “self-introductions” of natural enemies of arthropod pests and invasive plants. They conclude that these “self-introductions” might exceed the number of species introduced intentionally. These introductions have been facilitated by the usual factors: the general surge in international trade; lack of surveillance for species that are not associated with live plants or animals; inability to detect or intercept microorganisms; huge invasive host populations that allow rapid establishment of their accidentally introduced natural enemies; and lack of aggressive screening for pests already established. Examples cited include species introduced to the United States’ mainland and Hawai`i specifically.
The U.S. Capitol – one of the entities that can reflect our priorities in setting phytosanitary policy
As I point out often, altering human activities that facilitate invasion is a political process. So is amending international agreements that are not effective. We need to determine the cause of the failures of the existing institutions and act to rectify them. See my critiques of both the American and international phytosanitary system Fading Forests II and Fading Forests III (see links at the end of this blog) and my earlier blogs, especially this and this.
SOURCES
Fenn-Moltu, G., S. Ollier, O.K. Bates, A.M. Liebhold, H.F. Nahrung, D.S. Pureswaran, T. Yamanaka, C. Bertelsmeier. 2023. Global flows of insect transport and establishment: The role of biogeography, trade and regulations. Diversity and Distributions DOI: 10.1111/ddi.13772
Weber, D.C., A.E. Hajek, K.A. Hoelmer, U. Schaffner, P.G. Mason, R. Stouthamer, E.J. Talamas, M. Buffington, M.S. Hoddle and T. Haye. 2020. Unintentional Biological Control. Chapter for USDA Agriculture ResearchService. Invasive Insect biocontrol and Behavior Laboratory. https://www.ars.usda.gov/research/publications/?seqNo115=362852
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Clive Braser and others study Phytophthora species in their native habitats of Vietnam; which will become aggressive invaders in North America?
For years, one focus of this blog has been on scientists’ efforts to improve prevention of new introductions of forest pests. In earlier blogs, I summarized and commented on efforts by Mechet al. (2019) and Schultzet al. (2021), who extrapolate from insect-host relationships of pests already established in North America. [Full citations are presented at the end of this blog.] Both limited their analysis to insects; Mech et al. focused on those that attack conifers, Schultz et al. on those that attack single genera of angiosperms (hardwoods).
However, many of the most damaging agents are pathogens; for an indication, review the list under “invasive species” here. Indeed, Beckman et al. (2021) reported that only three non-native organisms pose serious threats to one or more of the 37 species of Pinus native to the U.S. All are pathogens: white pine blister rust (WPBR), pitch canker, and Phytophthora root rot (Phytophthora cinnamomi).
For this reason I welcome a study by Li et al. (2023), who used laboratory tests to evaluate the threat posed by more than 100 fungi associated with bark beetles. Since there are more than 6,000 species of bark and ambrosia beetles and they are commonly intercepted at the U.S. border, determining which should be priorities is important. Li et al. point out that the vast majority of such introductions have had minimal impacts. Two, however, have caused disastrous levels of damage: Dutch elm disease and laurel wilt disease.
Li et al. tested 111 fungi associated with 55 scolytine beetles from areas of Eurasia with latitudes and ecosystems analagous to those in the southeastern U.S. The beetles assessed included beetle species responsible for recent major tree mortality events in Eurasia: Dendroctonus species, Platypus koryoensis (Korean oak wilt), Platypus quercivorus (Japanese oak wilt) and Tomicus species.
The authors tested the fungi’s virulence on four species of trees native to the Southeast – two pines (Pinus taeda and P. elliottii var. elliottii), and two oaks(Quercus shumardii and Q. virginiana).
Li et al. found that none of 111 fungal associates caused a level of damage on these four hosts equal to Dutch elm disease on elms or laurel wilt disease on trees in the Lauraceae. Twenty-two of the fungi were minor pathogens – meaning they might cause damage under certain conditions or when loads of inoculum are large enough.
redbay trees killed in coastal Georgia by laurel wilt; photo by Scott Cameron
I think Li et al. set an extremely high bar for “serious” damage. Surely we wish to prevent introduction of pathogens that cause damage at a lower level than the catastrophes to which these two diseases have exposed a genus (elms) and a family (Lauraceae)! Still, the scientific approach used here is a step toward addressing pathogens. These agents of tree mortality are addressed much less frequently than insects. I hope that scientists will continue to test the virulence of these fungi on some of the thousands of other species that make up the forests of the United States, or at least the dominant species in each ecosystem.
It is discouraging that Raffaet al. (2023) found none of four approaches to predicting a new pest’s impact to be adequate by itself. Instead, they outlined the relative strengths and weaknesses of each approach and the circumstances in which they might offer useful information. I am particularly glad that they have included pathogens, not just insects. The four approaches they review are:
(1) pest status of the organism in its native or previously invaded regions;
(2) statistical patterns of traits and gene sequences associated with high-impact pests;
(3) sentinel plantings to expose trees to novel pests; and
(4) laboratory tests of detached plant parts or seedlings under controlled conditions.
They emphasize that too little information exists regarding pathogens to predict which microbes will become damaging pathogens when introduced to naïve hosts in new ecosystems. See the article, especially Figure 4, for their assessment of the strengths each of the several approaches.
Raffa et al. raise important questions about both the science and equity issues surrounding invasive species. As regards scientific issues, they ask, first, whether it will ever be possible to predict how each unique biotic system will respond to introduction of a new species. Second, they ask how assessors should interpret negative data? In the context of equity and political power, they ask who should make decisions about whether to act?
In my blog I expressed concern about finding that most introduced forest insects are first detected in urban areas whereas introduced pathogens are more commonly detected in forests. I hope scientists will redouble efforts to improve methods for earlier detection of pathogens. Enrico Bonello at Ohio State and others report that spectral-based tools can detect pathogen-infected plants, including trees.
Japanese cherry trees burned on the Washington D.C. mall because infested by scale; on order of Charles Marlatt
International trade is considered the single most important pathway for unintentional introductions of insects. Updated figures remind us about the stupendous amounts of goods being moved internationally. According to Weber et al., international shipping moves ~133 million TEU containers per year between countries, the majority between continents. Four times this number move within regions via coastal shipping. On top of that, four billion passenger trips take place by air every year. Air freight carries another ~220 million tons of goods; while this is a tiny fraction of the weight shipped by boat, the packages are delivered in less than a day – greatly increasing the likelihood that any unwanted living organisms will survive the trip. The U.S. also imports large numbers of live plants – although getting accurate numbers is a challenge. MacLachlan et al. (2022) report 5 billion plants imported in 2021, but the USDA APHIS annual report for FY22 puts the number at less than half that figure: 2.2 billion plant units.
Given the high volume of incoming goods, Weber et al. advocate improved surveillance (including analysis of corresponding interceptions) of those pathways that are particularly likely to result in non-native species’ invasions, e.g. live plants, raw lumber(including wood packaging), and bulk commodities e.g. quarried rock. Isitt et al. and Fenn-Moltu et al. concur that investigators should focus on the trade volumes of goods that are likely to transport plant pests – in their cases, plant imports.
The importance of the plant trade as a pathway of introduction for has been understood for at least a century – as witnessed by the introductions of chestnut blight DMF and white pine blister rust, DMF and articles by Charles Marlatt. A decade ago, Liebhold et al. (2012) calculated that the approach rate of pests on imported plants was 12% — more than 100 times higher than the 0.1% approach rate found by Haack et al. (2014) for wood packaging.
Since plant-insect interactions are the foundation of food webs, changes to a region’s flora will have repercussions throughout ecosystems, including insect fauna. See findings by teams led by Doug Tallamy and Sara Lalk; and a chapter in the new forest entomology text written by Bohlmann, and Krokene (citation at end of blog under Allison, Paine, Slippers, and Wingfield). Sandy Liebhold and Aymeric Bonnamour also addressed explicitly links between introductions of non-native plant and insect species. Weber et al. call this phenomenon the “receptive bridgehead effect”: a non-native plant growing prolifically in a new ecosystem provides a suitable host for an organism that feeds on that host, raising the chance for its establishment.
Recent studies confirm the importance of the “receptive bridgehead effect”. Isitt and colleagues found that the large numbers of introduced European insect species – all taxa, not just phytophagous insects – established in North America and Australia/New Zealand were best explained by the numbers of European plants introduced to these regions – in other words, the most important driver appears to be the diversity of non-native plants.
The presence of European plants in North America and Australia/New Zealand promoted establishment of European insects in two ways. First, these high-volume imports increased the propagule pressure of insects associated with this trade. Live plant imports might have facilitated the establishment of ~70% of damaging non-native forest insects in North America. Second, naturalization of introduced European plants provided a landscape replete with suitable hosts. This is especially obvious in Australia/New Zealand, which have unique floras. In Australia, nearly 90% of non-native pest insects are associated with non-native plants. Those non-native insects that do feed on native plants are more likely to be polyphagous.
Amur honeysuckle – one of the hundreds of Asian plants invading North American ecosystems; via Flickr
I hope U.S. phytosanitary officials apply these lessons. Temperate Asia is the source of more non-native plants established in both North America and Australia/New Zealand than is Europe. Already, many insects from Asia have invaded the U.S. The logicof the “receptive bridgehead effect” points to prioritizing efforts to prevent even more Asian insects from reaching our shores!
Fenn-Moltu et al. sought to elucidate which mechanisms facilitate species’ success during the transport and introduction/establishment stages of bioinvasion. They studied the transport stage by analyzing border interceptions of insects from 227 countries by Canada, mainland U.S., Hawai`i, Japan, New Zealand, Great Britain, and South Africa over the 60 year period 1960 – 2019. They studied establishment by analyzing attributes of 2,076 insect species recorded as established after 1960 in the above areas plus Australia (North America was treated as a single unit comprised of the continental U.S. and Canada).
The number of species transported increased with higher Gross National Income in the source country. The number of species transported decreased with geographic distance. They suggest that fewer insects survive longer journeys, but say additional information is needed to verify this as the cause. The number of species transported was not affected by species richness in the native region.
More species established when introduced to a country in the same biogeographic region. They were not surprised that environmental similarity between source and destination apparently strongly affected establishment success. The number of species established was not affected by species richness in the native region. For example, the greatest number of established species originated from the Western and Eastern Palearctic regions, which together comprise only the fifth-largest pool of native insect species.
Gaps Despite Above Studies
As I noted at the beginning, most of the studies examining current levels of pests transported on imported plants have been limited to insects. This is unfortunate given the impact of introduced pathogens (again, review the list damaging organisms under “invasive species” here).
In addition, most studies analyzing the pest risk associated with plant imports use port inspection data – which are not reliable indicators of the pest approach rate. The unsuitability of port inspection data was explained by Liebhold et al. in 2012 and Fenn-Moltu et al. a decade later – as well as Haack et al. 2014 (as the data pertain to wood packaging). Fenn-Moltu et al. note that inspection agencies often (and rightly!) target high-risk sources/commodities, so the records are biased. Other problems might arise from differences in import volume, production practices, and differences in records that identify organism only to genus level rather than species. Fenn-Moltu et al. call for relying on randomized, statistically sound inspection systems; one such example is USDA’s Agriculture Quarantine Inspection System (AQIM). Under AQIM, incoming shipments are randomly selected and put through more thorough inspections to produce statistically based estimates of approach rates, defined as the percent of inspected shipments found to be infested with potential pests (Liebhold et al. 2012). I ask why scientists who are aware of this issue have not obtained AQIM data for pests associated with plant imports. Plant imports have been included in the AQIM system since 2008. Have they not been able to persuade APHIS to provide these data? Or are these data available for only limited types of imported plants? Too narrow a focus would create a different source of potential bias.
Both Isitt et al. and Fenn-Moltu et al. list factors not addressed and other caveats of which we should be aware when extrapolating from their findings.
Beckman, E., Meyer, A., Pivorunas, D., Hoban, S., & Westwood, M. (2021). Conservation Gap Analysis of Native U.S. Pines. Lisle, IL: The Morton Arboretum.
Fenn-Moltu, G., S. Ollier, O.K. Bates, A.M. Liebhold, H.F. Nahrung, D.S. Pureswaran, T. Yamanaka, C. Bertelsmeier. 2023. Global flows of insect transport and establishment: The role of biogeography, trade and regulations. Diversity and Distributions DOI: 10.1111/ddi.13772
Hoddle. M.S. 2023. A new paradigm: proactive biological control of invasive insect pests. BioControl https://doi.org/10.1007/s10526-023-10206-5
Isitt, R., A.M. Liebhold, R.M. Turner, A. Battisti, C. Bertelsmeier, R. Blake, E.G. Brockerhoff, S.B. Heard, P. Krokene, B. Økland, H. Nahrung, D. Rassati, A. Roques, T. Yamanaka, D.S. Pureswaran. 2023. Drivers of asymmetrical insect invasions between three world regions. bioRxiv preprint doi: https://doi.org/q0.1101/2023.01.13.523858
Li, Y., C. Bateman, J. Skelton, B. Wang, A. Black, Y-T Huang, A. Gonzalez, M.A. Jusino, Z.J. Nolen, S. Freemen, Z. Mendel, C-Y Chen, H-F Li, M. Kolarik, M. Knizek, J-H. Park, W. Sittichaya, T-H Pham, S. Ito, M. Torii, L. Gao, A.J. Johnson, M. Lu, J. Sun, Z. Zhang, D.C. Adams, J. Hulcr. 2022. Pre-invasion assessment of exotic bark beetle-vectored fungi to detect tree-killing pathogens. Phytopathology Vol 112 No. 2 February 2022
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. www.frontiersinecology.org
Liebhold, A.M., T. Yamanaka, A. Roques, S. August, S.L. Chown, E.G. Brockerhoff and P. Pyšek. 2018. Plant diversity drives global patterns of insect invasions. Sci Rep8, 12095 (2018). https://doi.org/10.1038/s41598-018-30605-4
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Mech, A.M., K.A. Thomas, T.D. Marsico, D.A. Herms, C.R. Allen, M.P. Ayres, K.J. K. Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, and P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol Evol. 2019 Nov; 9(21): 12216–12230.
Raffa, K.F., E.G. Brockerhoff, J-C. Gregoirem R.C. Hamelin, A.M. Liebhold, A. Santini, R.C. Venette, and M.J. Wingfield. 2023. Approaches to Forecasting Damage by Invasive Forest Insects and Pathogens: A Cross-Assessment. Bioscience Vol. 73, No. 2. February 2023.
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions.
Weber, D.C., A.E. Hajek, K.A. Hoelmer, U. Schaffner, P.G. Mason, R. Stouthamer, E.J. Talamas, M. Buffington, M.S. Hoddle and T. Haye. 2020. Unintentional Biological Control. Chapter for USDA Agriculture ResearchService. Invasive Insect biocontrol and Behavior Laboratory. https://www.ars.usda.gov/research/publications/?seqNo115=362852
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
As we know, survival of North American species of ash (Fraxinus spp.) is threatened by the emerald ash borer (EAB). DMF Sadof, McCullough, and Ginzel (full citation at end of the blog) hope to prevent demise of another ~ 135 million urban ash trees by 2050 bycountering persistent myths that have hindered adoption of effective protective measures. As they note, USDA APHIS has dropped regulations that had been intended to slow the EAB’s spread – which I concede were not very effective.
Protecting urban ash trees now falls to municipalities, states, their leaders and citizens, non-governmental organizations, and tree care professionals. If they apply knowledge gained since the detection of EAB 20 years ago – and are not paralyzed by myths – they can successfully manage EAB populations and protect their town’s ash trees. [I have also blogged about efforts to breed ash trees resistant to EAB.]
Since some studies have found that “myth-busting” is not effective, perhaps people advocating for EAB control should avoid mentioning the myths per se and instead emphasize the science supporting the proposed actions.
Sadof, McCullough, and Ginzel first review aspects of the biology of ash and EAB that are relevant to arborists and pest management specialists:
Adult EAB beetles feed on tree leaves for a couple of weeks from mid-May through June. This maturation period provides a 2–3 week opportunity to kill the leaf-feeding beetles with systemic insecticides before any eggs are laid.
Once eggs hatch, the first stage larvae immediately move into the phloem (inner bark) and cambium tissue, where they begin feeding. Systemic insecticides rarely enter the phloem, so they kill few larvae during this stage.
Detection of early stages of invasion is hampered by several factors, including beetles’ initial colonization of branches in the upper canopy; initially minimal effect on healthy ash trees; and the frequency of two-year life cycles when beetle densities are low. However, it is important to detect and treat these early infestations because EAB populations increase, tree health declines to eventual death.
Detection efforts should target the ash trees most likely to be infested early in the invasion: stressed trees, preferred species (especially green ash), trees growing in the open in parks, along roadsides or surrounded by impervious surfaces. Authorities can take advantage of the attractiveness of stressed trees by establishing “trap trees” to attract EAB adults. Beetles that feed on the “trap trees” can be killed by systemic insecticides. Or the trees can be removed and chipped to kill eggs and larvae before they can emerge. Sadof, McCullough, and Ginzel say trap trees are effective in slowing spread of new infestations when most ash trees remain healthy. Once EAB densities build and many trees are stressed by larval feeding, volatile (airborne) compounds released by girdled trees no longer attract the beetles.
Woodpecker holes in branches of the upper canopy are often the first evidence of EAB invasion in an area.
Even in late stages of the invasion, when most ash trees that were not protected with systemic insecticides are dead, EAB populations persist and continue to colonize and kill available ash trees, including some as small as >2.5 cm in diameter.
Myth: There Is No Point in Trying to Protect Ash Trees—
EAB Will Eventually Kill Them Anyway
Answer:
When the EAB was first detected in 2002, control measures were limited in number and efficacy. In the 20 intervening years, scientist have learned much about EAB biology and ash physiology. Insecticide chemistry and application methods have improved. Currently recommended strategies are based on long-term field studies. More effective insecticides have been developed. Emamectin benzoate is particularly efficient, including the fact that it needs to be applied only every third year. Managers must pay attention to the application protocols, including appropriate dose (i.e., the amount of insecticide product applied); spacing injection ports around the trunk to ensure that the xylem will transport the chemical to leaves throughout the canopy; and conduct injections in spring after bud break.
Myth: Wounds From Drilling Trees to Inject Systemic Insecticides Injure Trees
Answer:
In the early years, trunk injections sometimes caused substantial injury to trees. Refinement of delivery devices and reductions in the pressure at which insecticides are injected have virtually eliminated these issues. Staff must be properly trained in use of the equipment.
demonstration of injecting pesticide into ash tree; photo by F.T. Campbell
Myth: Using Systemic Insecticides to Protect Ash Trees Harms
Non-target Species and the Environment
Answer:
Sadof, McCullough, and Ginzel point out that continent-wide loss of a tree genus is likely to adversely affect the more than 200 species of native arthropods that are specialists on ash. On the other hand, systemic insecticides are unlikely to harm beneficial natural enemies of EAB, including parasitoid wasps, predatory insects, or woodpeckers. First, the insecticides are contained within the tree’s tissues; they do not kill insects on contact. Second, parasitoids and predators avoid dead beetles. Honeydew excreted by sucking insects might contain sufficient insecticide residue to harm parasitoids — if the tree is heavily infested. However, these insects are rapidly killed by these insecticides if they are applied at the optimal time (early to mid-spring). Proper timing of application greatly reduces the potential for tainted honeydew to accumulate on infested trees. Furthermore, in cities there are few populations of natural enemies of sucking insects.
Most concern is focused on pollinators. Ash trees flower early, before leaves expand. It is reassuring that protocols instruct that the systemic insecticides be applied after bud break — typically after pollen has been shed. I do find it disturbing that apparently there have been no published studies of insecticide concentration in ash pollen.
Myth: It Costs Too Much to Protect Ash Trees
Answer:
Sadof, McCullough, and Ginzel review the several studies and methods developed to estimate the value of urban ash trees – both individually and over a wider area. The value is based on the individual tree’s location, health, and structural condition. These economic studies have consistently shown that it costs less to protect ash trees from EAB with insecticide treatments than to remove ash trees — either proactively or when they decline and die.
Even delaying tree mortality – short of preventing it completely – is worthwhile because it allows municipalities to incorporate tree removal into the budget, rather than be suddenly confronted by large expense that they had not planned for.
Sadof, McCullough, and Ginzel recommend treating ash within a significant area as being most efficient. This approach reduces overall costs and slows rates of ash mortality locally – even for trees that are not treated. In some cases, treating as few as 11% of ash trees slowed the overall rate of ash decline.
An important in comparing costs of treatment to costs of replacement is the high mortality rate of newly planted urban trees: up to two-thirds die shortly after planting. This means that it takes decades to replace a mature tree canopy and the environmental benefits the canopy provides. Sadof, McCullough, and Ginzel conclude that protecting ash trees from EAB has clear positive effects for both the urban forest canopy – and its environmental services – and municipal forestry budgets.
Sadof, McCullough, and Ginzel then outline a viable Integrated Pest Management (IPM) framework that incorporates use of systemic insecticides to protect ash trees from EAB.
1. Define the problem and identify management objectives
Inventory urban trees before EAB is detected. The inventories should identify priority trees based on size (diameter at breast height), tree condition, and suitability of the site where the tree is growing. Focus detection surveillance on green ash trees, especially those in parks, parking lots, and along roads — sites that are sunlit (open) and likely to cause stress to the trees.
2. Monitor and assess the local EAB population to determine when a treatment program should be initiated. Treatment must wait until there is evidence that EAB is presentbut should not then be delayed, since it should begin while the trees’ vascular systems are still sufficiently healthy to carry the insecticide to branches and leaves. This requires regular inspections of ash trees for visible signs of EAB infestation. Efficiency is improved by focusing on high-risk trees (see above) and noticing woodpecker holes on upper portions of the trunk. Consider debarking symptomatic trees or establishing “trap tree” networks.
3. Identify and gather resources needed to implement an insecticide treatment program. Web-based calculators guide budget decisions based on the municipality’s tree inventory and local costs of treatments. Treating one-third of trees annually with emamectin benzoate can save money while maximizing the number of trees protected. Training city forestry staff in trunk injection methods is cheaper than hiring contractors and ensures better treatment quality and efficiency.
downy woodpecker; photo by Steven Bellovin, Columbia University
4. Incorporate multiple tactics to protect tree health and control EAB.
Ensure trees are actively transpiring when injecting the systemic insecticides; this might requireirrigation. Encourage parasitoids and woodpecker foraging on untreated trees. In areas where ash trees are closely spaced, consider an area-wide urban SLAM program. In this strategy, treating a proportion of ash trees at two-year intervals reduces EAB eggs and overall EAB populations. Non-treated trees with EAB larvae might support parasitoid biocontrol populations whose offspring can attack EAB larvae on previously treated ash trees as the emamectin benzoate concentration wanes.
Sadof, McCullough, and Ginzel also suggest establishing a citizen monitoring program to both reduce costs and build community support for ash management. Community participation has been particularly effective when professionals take appropriate and timely action in response to volunteers’ findings.
SOURCE
Sadof, C.S., D.G. McCullough, and M.D. Ginzel. 2023. Urban ash management and emerald ash borer (Coleoptera: Buprestidae): facts, myths, and an operational synthesis. Journal of Integrated Pest Management, 2023, Vol. 14, No. 1 https://doi.org/10.1093/jipm/pmad012
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
beech leaf disease symptoms; photo courtesy of Jennifer Koch, USFS
As we know, beech leaf disease (BLD) has spread rapidly in the decade since its discovery in northeast Ohio. It has been detected as far east as the Maine coast, as far south as northern Virginia, as far north as southern Ontario, and as far west as eastern Michigan and northern Indiana. It has been found in 12 states.
BLD is associated with a nematode, Litylenchus crenatae subsp. mccannii (Lcm), although whether this is the sole causal agent is not yet clear.
BLD’s North American host, American beech (Fagus grandifolia),is an important native deciduous hardwood species. It plays important roles in nutrient cycling, erosion control, and carbon storage and sequestration in forests. Wildlife species depend on the trees’ canopies and especially cavities blog for nesting sites, shelter, and nutritious nuts. American beech – with sugar maple (Acer saccharum) and yellow birch (Betula alleghaniensis) – dominate the northern hardwood ecosystem of northeastern United States and southeastern Canada. These forests occupy a huge area; in just New England and New York they occupy 20 million acres (Leak, Yamasaki and Holleran. 2014; full citation at end of blog).
Beech leaf disease also affects European beech, (F. sylvatica), Chinese beech (F. engleriana), and Oriental beech (F. orientalis) planted in North America. The disease has not yet been detected in Asia or Europe. Japanese beech (F. crenata) sporadically display symptomatic leaves, but the disease has not been reported there.
Scientists working to understand the disease, how it spreads, and its ecological impact confer every other month. The next time is in early December.
Paulo Vieira, of the USDA Agriculture Research Service, leads one group seeking to better understand how the disease infects its host. They published a new study (see full citation at end of blog) examining how the nematode provokes changes in the cells of the trees’ leaves. As they point out, leaves are plants’ primary organs for photosynthesis – hence providing energy for growth. The leaf is composed of a several cell types organized into different tissues with specific function related to photosynthesis, gas exchange, and/or the transportation of water and nutrients. Thus, changes in leaf morphology affect the normal functioning of the leaf and therefore the tree’s growth and survival.
Vieira et al. found that:
The BLD nematode enters the leaf bud as it forms in late summer. In early autumn, all nematode developmental stages were found in the buds, including eggs at various stages of embryonic development, juveniles, and adults. Adult males were found in fewer than 20% of the buds, suggesting that the nematode can reproduce asexually.
Feeding by the BLD nematode induces abnormal and extensive cell proliferation, resulting in a significant increase of the number of cell layers inside the leaf. These changes improve the nutrition that the leaves provide to the nematode. However, the BLD-induced distortions of the bud persist as the leaf grows. Symptomatic leaf “banding” results. These areas have a proliferation of abnormally large and irregularly shaped cells with more chloroplasts. Intercellular spaces are also larger; this is where the nematodes are found. in. (The publication has dramatic photographs.)
Sites damaged by nematodes are a major resource for metabolites needed for plant performance. So their damage imposes a considerable drain.
Colonization of roots by ectomycorrhizal fungal is also reduced in severely diseased trees.
Immature female nematodes are the principal winter survivors. However, many die, making it difficult to culture nematodes in the spring. The nematodes reproduce during the growing season. Buildup of nematode numbers makes culturing easier, so facilitating confirmation of the disease’s presence.
Nematodes can migrate along the stem to other leaves, thus spreading the infection.
Vieira et al. tell us fascinating facts about the nematode. The BLD nematode, Litylenchus crenatae subsp. mccannii (Lcm) is now considered one of the top ten most important plant-parasitic nematodes in the United States. To date, species of this genus have been found only in Japan and New Zealand. The species L. crenatae was first described from Japan. A second species — L. coprosma – was detected in 2012 in New Zealand in association with small chlorotic patches on leaves of two native plants in the Coprosma genus.
Litylenchus belongs to the family Anguinidae. Several species in the family are designated quarantine pests because they cause economically significant damage to food and ornamental corps, including grains (wheat, barley, rice) and potatoes. Anguinidae nematodes often parasitize aerial parts of the hosts (e.g., leaves, stems, inflorescences, seeds); less frequently they infest roots. They can migrate along the host tissue surfaces in water films. Their host ranges vary from broad to narrow. Other Anguinidae nematodes apparently share the ability to manipulate the host’s cellular machinery, which often results in the induction of cell hyperplasia [the enlargement of an organ or tissue caused by an increase in the reproduction rate of its cells], and hypertrophy [increase and growth of cells] of the tissues on which they feed.
healthy beech leaves; F.T. Campbell
Vieira et al. assert that the rapid spread of Litylenchus crenatae subsp. mccannii – combined with the apparent lack of resistance in native beech trees – suggests that this nematode was recently introduced to North America. Furthermore, the ability of this subspecies to change the host’s cell cycle machinery supports the link between the presence of the nematode and the disease.
The mechanisms by which nematodes change host-plant cells are unknown. I hope that scientists will pursue these questions. Perhaps the nematode family’s threat to grains and other food crops will prompt funding for such work. Unfortunately, I don’t think the threat to an ecologically-important native tree species will have the same power.
SOURCES
Leak, W.B, M. Yamasaki and R. Holleran. 2014 Silvicultural Guide for Northern Hardwoods in the Northeast. United States Department of Ariculture Forest Service Northern Research Station. General Technical Report NRS-132. April 2014.
Vieira P, M.R. Kantor MR, A. Jansen, Z.A. Handoo, J.D. Eisenback. (2023) Cellular insights of beech leaf disease reveal abnormal ectopic cell division of symptomatic interveinal leaf areas. PLoS ONE October 5, 2023. 18(10) https://doi.org/10.1371/pone.0292588
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
In October 2018 I posted a blog on the decline of global insect numbers and biodiversity.
This month a Washington Post columnist, Michael J. Corin, published a piece decrying people’s use of “bug zappers” [full citation at end of this blog] in an effort to prevent biting insects from ruining their evenings.
After quoting some of the manufacturers’ advertising claims, Corin point to the scientific consensus that these zappers don’t kill mosquitoes. In some studies, the zappers killed tens of thousands of insects, but far fewer than 1% were mosquitoes. A study by Iowa State University (citation at end of blog) that estimated even a fraction of the bug zappers sold in the United States kill more than 70 billion insects annually.
Corin even says, “bug zappers make it more likely you’ll be bitten by mosquitoes while sitting in your backyard.” Any mosquitoes drawn to the vicinity of a bug zapper will redirect their attention to the proximal warmblooded mammals — usually humans.
This information is important to us because bug zappers are exceptional killers of (other) insects. Corin cites a study by the University of Delaware (Frick and Tallamy, full citation below) in which the zappers caught nearly 14,000 insects over a summer. Roughly half the catch — 6,670 insects — were harmless aquatic species from nearby rivers and streams, fish food in the aquatic food chain. Many of the others were parasitic wasps and beetles that naturally prey on mosquitoes.
caddisfly (one of groups often killed by bug zappers, according to Doug Tallamy) photo by Anita Gould via Flickr
So, these traps are exacerbating the “insect apocalypse” and undermining biocontrol programs!
These studies were published in the middle-1990s – nearly 30 years ago. Dr. Douglas Tallamy of the University of Delaware told me that there are now traps that use baits (octanol and CO2). Carbon dioxide and octenol (a derivative of mammalian body odor) are known to attract biting insects, including mosquitoes. However, these traps cost up to $500 and do not sell well. The standard zapper still only catches non-targets.
A study by Kim et al. found that neither the Stinger Electric Zapper nor the Mosquito Deleto works effectively.
Apparently most bug zappers are not working as advertised. Why, then, are they still marketed using false claims? The Federal Trade Commission is supposed to investigate misleading advertising claims and take legal action against manufacturers who don’t correct them. Corin says the agency suggested to him that the public should submit any complaints through the agency’s website.
Should we not provide information to the FTC and urge them to take action? Do you have information – or access to research capabilities – to support such an effort?
Corin provides the usual advice to minimize mosquitoes around your house:
1) eliminate standing water – in which mosquitoes breed. Or install a “Bucket of Doom” (a design developed by the Centers for Disease Control). Fill a 5-gallon bucket with water and add leaf litter or straw.Mosquitoes love to lay eggs here. Add granules of Bacillus thuringiensis to kill the mosquito larvae. Some commercial versions are Ovi-Catch AGO trap sold by Catchmaster or the “GAT” trap sold by Biogents.
2) wear long sleeves and pants. Add repellent – especially those containing DEET.
Frick, T.B. and D.W. Tallamy. 1996. Density and Diversity of Nontarget Insects Killed by Suburban Electric Insect Traps. Ent. News Vol 107, No. 2, March & April 1996
Kim, J. et al. 2002. A study comparing efficiency of insect capture between Stinger electric zapper and Mosquito-deleto at varying locations and heights in northern Michigan. https://deepblue.lib.umich.edu/handle/2027.42/54969
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
tanoak trees in southern Oregon killed by sudden oak death; photo by Oregon Department of Forestry; this pathogen is not mentioned by USFS RPA report
In August the USDA Forest Service published the agency’s 2020 assessment of the future of America’s forests under the auspices of the Resources Planning Act. [See United States Department of Agriculture Forest Service Future of America’s Forests and Rangelands, full citation at the end of the blog.] To my amazement, this report is the first in the series (which are published every ten years) to address disturbance agents, specifically invasive species. In 2023! Worse, I think its coverage of the threat does not reflect the true state of affairs – as documented by Forest Service scientists among others.
This is most unfortunate because policy-makers presumably rely on this report when considering which threats to focus on.
Here I discuss some of the USFS RPA report and what other authors say about the same topics.
The RPA Report’s Principle Foci: Extent of the Forest and Carbon Sequestration
The USFS RPA report informs us that America’s forested area will probably decrease 1- 2% over the next 50 years (from 635.3 million acres to between 619 and 627 million acres), due largely to conversion to other uses. This decline in extent, plus trees’ aging and increases in disturbance will result in a slow-down in carbon sequestration by forests. In fact, if demand for wood products is high, or land conversion to other uses proceeds apace, U.S. forest ecosystems are projected to become a net source of atmospheric CO2 by 2070.
Eastern forests sequester the majority of U.S. forest carbon stocks. These forests are expected to continue aging – thereby increasing their carbon storage. Yet we know that these forests have suffered the greatest impact from non-native pests.
I don’t understand why the USFS RPA report does not explicitly address the implications of non-native pests. In 2019, Songlin Fei and three USFS research scientists did address this topic. Fei et al. estimated that tree mortality due to the 15 most damaging introduced pest species have resulted in releases of an additional 5.53 terragrams of carbon per year. Fei and colleagues conceded this is probably an underestimate. They say that annual levels of biomass loss are virtually certain to increase because current pests are still spreading to new host ranges (as demonstrated by detection of the emerald ash borer in Oregon). Also, infestations in already-invaded ranges will intensify, and additional pests will be introduced (for example, beech leaf disease).
I see this importance of eastern forests in sequestering carbon as one more reason to expand efforts to protect them from new pest introductions, and the spread of those already in the country, etc.
A second issue is the role of non-native tree species in supporting the structure and ecological functions of forests. Ariel Lugo and colleagues report that 18.8 million acres (7.6 million ha, or 2.8% of the forest area in the continental U.S.) is occupied by non-native tree species. (I know of no overall estimate for all invasive plants.) They found that non-native tree species constitute 12–23% (!) of the basal area of those forest stands in which they occur.
Norway maple (Acer platanoides); one of the most widespread invasive species in the East. Photo by Hermann Falkner via Flickr
Lugo and colleagues confine their analysis of ecosystem impacts to carbon sequestration. They found that the contribution of non-native trees to carbon storage is not significant at the national level. In the forests of the continental states (lower 48 states), these trees provide 10% of the total carbon storage in the forest plots where they occur. (While Lugo and colleagues state that the proportion of live tree biomass made up of non-native tree species varies greatly among ecological subregions, they do not provide examples of areas on the continent where their biomass – and contribution to carbon storage — is greater than this average.) In contrast, on Hawai`i, non-native tree species provide an estimated 29% of live tree carbon storage. On Puerto Rico, they provide an even higher proportion: 36%.
Brazilian pepper (Schinus terebinthifolius) – widespread invasive in Hawai`i and Florida; early stage invasive in Puerto Rico. Photo by Javier Alexandro via Flickr
In the future, non-native trees will play an even bigger role. Since tree invasions on the continent are expanding at ~500,000 acres (202,343 ha) per year, it is not surprising that non-native species’ saplings provide 19% of the total carbon storage for that size of trees in the lower 48 states (Lugo et al.).
Forming a More Complete Picture: Biodiversity, Disturbance, and Combining Data.
The USFS RPA report has a chapter on biodiversity. However, the chapter does not discuss historic or future diversity of tree species within biomes, nor the genetic diversity within tree species.
Treatment of Invasive Species
The USFS 2020 RPA report is the first to include a chapter on disturbance, including invasive species. I applaud its inclusion while wondering why they have included it only now? Why is the coverage so minimal? I think these lapses undercut the report’s purpose. The RPA is supposed to inform decision-makers and stakeholders about the status, trends, and projected future of renewable natural resources and related economic sectors for which USFS has management responsibilities. These include: forests, forest products, rangelands, water, biological diversity, and outdoor recreation. The report also has not met its claim to “capitalize on” areas where the USFS has research capacity. One excuse might be that several important publications have appeared after the cut-off date for the assessment (2020). Still, the report’s authors cite some of the evaluations that were in preparation as of 2020, e.g., Poland et al.
I suggest also that it would be helpful to integrate data from other agencies, especially the invasive species database compiled by the U.S. Geological Survey, into the RPA. For example, the USGS lists just over 4,000 non-native plant species in the continental U.S. (defined as the lower 48 plus Alaska). On Hawai`i, the USGS lists 530 non-native plant species as widespread. Caveat: many of the species included in these lists probably coexist with the native plants and make up minor components of the plant community.
Specifically: Invading Plants
The USFS RPA report gives much more attention to invasive plants than non-native insects and pathogens. The report relies on the findings of Oswalt et al., who based their data on forested plots sampled by the Forest Inventory and Analysis (FIA) program. (The RPA also reports on invasive plants detected on rangelands, primarily grasslands.) Oswalt et al. found that 39% of FIA plots nationwide contained at least one plant species that the FIA protocol considers to be invasive and monitors. The highest intensity of plant invasions is in Hawai`i – 70% of the plots are invaded. The second-greatest intensity is in the eastern forests: 46%. However, the map showing which plots were inventoried for invasive plants makes clear how incomplete these data are – a situation I had not realized previously.
I appreciate that the USFS RPA report mentions that propagule pressure is an important factor in plant invasions. This aspect has often been left out in past analyses. I also appreciate the statement that international trade in plants for ornamental horticulture will probably lead to additional introductions in the future. Third, I concur with the report’s conclusions that once forest land is invaded, it is unlikely to become un-invaded. Invasive plant management in forests often results in one non-native species being replaced by another. In sum, the report envisions a future in which plant invasion rates are likely to increase on forest land.
If you wish to learn more about invasive plant presence and impacts, see the discussion of invasive plants in Poland et al., my blogs based on the work by Doug Tallamy, and several other of my blogs compiled under the category “invasive plants” on this website.
I believe all sources expect that the area invaded by non-native plant species, and the intensity of existing invasions, will increase in the future.
The USFS RPA links these invasions to expansion of the “wildland-urban interface” (“WUI”). These areas increased rapidly before 2010. At that time, they occupied 14% of forest land. The report published in 2023 did not assess their future expansion over the period 2020 to 2070. However, it did project increased fragmentation in many regions, especially in the RPA Western and Southeastern regions. Since “fragmentation” is very similar to wildland-urban interfaces, the report seems implicitly to project more widespread plant invasions in the future.
plant invasions facilitated by fragmentation; northern Virginia; photo by F.T. Campbell
Specifically: Insects and Pathogens
The USFS RPA report on insects and pathogens is brief and contains puzzling errors and gaps. It says that the tree canopy area affected by both native and non-native mortality-causing agents has been consistently large over the three most recent five-year FIA assessment periods. It notes that individual insects or diseases have extirpated entire tree species or genera and fundamentally altered forests across broad regions. Examples cited are chestnut blight and emerald ash borer.
The USFS RPA report warns that pest-related mortality might be underreported in the South, masked by more intense management cycles and higher rates of tree growth and decay. On the other hand, the report asserts that pest-related mortality is probably overrepresented in the Northern Region in the 2002 – 2006 period because surveyors drew polygons to encompass large areas affected by EAB and balsam woolly adelgid (Adelges piceae) infestations. The latter puzzles me; I think it is probably an error, and should have referred to hemlock woolly adegid, A. tsugae. Documented mortality has generally been much more widespread from insects than diseases, e.g., bark beetles, including several native ones, across all regions and over time, especially in the West – where the most significant morality agents are several native beetles. The USFS RPA report mentions that the Northern Region has been particularly affected by non-native pests, including EAB, HWA, BWA, beech bark disease, and oak wilt. It mentions that Hawai`i has also suffered substantial impacts from rapid ʻōhiʻa death.
Defoliating insects have affected relatively consistent area over time. This area usually equaled or exceeded the area affected by the mortality agents. Principal non-native defoliators in the Northern Region have been the spongy moth (Lymantria dispar); larch casebearer (Coleophora laricella); and winter moth (Operophtera brumata). In the South they list the spongy moth.
More disturbing to me is the USFS RPA report’s conclusion that the future impact of forest insects is highly uncertain. The authorsblame the complexity of interactions among changing climate, those changes’ effects on insect and tree species’ distributions, and overall forest health. Also, they name uncertainty about which new non-native species will be introduced to the United States. I appreciate the report’s avoidance of blanket statements regarding the effects of climate change. However, other studies – e.g., Poland et al. – have incorporated these complexities while still offering conclusions about a number of currently established non-native pests. Finally, I am particularly dismayed that the USFS RPA does not provide analysis of any forest pathogens beyond the single mention of a few.
I am confused as to why the USFS RPA report makes no mention of Project CAPTURE (Conservation Assessment and Prioritization of Forest Trees Under Risk of Extirpation). This is a multi-partner effort to prioritize U.S. tree species for conservation actions based on invasive pests’ threats and the trees’ ability to adapt to them. Several USFS units participated, including the Southern Research Station, the Eastern Forest Environmental Threat Assessment Center, and the Forest Health Protection program. The findings were published in 2019. See here. Lead scientist Kevin Potter was one of the authors of the RPA’s chapter on disturbance.
redbay (Persea borbonia) trees in Georgia killed by laurel wilt; photo by Scott Cameron. Redbay is ranked by Project CAPTURE as 5th most severely at risk due to a non-native pest
“Project CAPTURE” provided useful summaries of non-native pests’ impacts, including the facts that
54% of the tree species on the continent are infested by one or more non-native insect or pathogen;
nearly 70% of the host/agent combinations involve angiosperm (broadleaf) species, 30% gymnosperms (e.g., conifers). When considering only non-native pests, pests attacking angiosperms had greater average severity.
Disease impacts are more severe, on average, than insect pests. Wood-borers are more damaging than other types of insect pests.
Non-native agents have, on average, considerably more severe impacts than native pests.
Project CAPTURE also ranked priority tree species based on the threat from non-native pests (Potter et al., 2019). Tree families at the highest risk to non-native pests are: a) Fagaceae (oaks, tanoaks, chestnuts, beech), b) Sapindaceae (soapberry family; includes maples, Aesculus (buckeye, horsechestnut); c) in some cases, Pinaceae (pines); d) Salicaceae (willows, poplars, aspens); e) Ulmaceae (elms) and f) Oleaceae (includes Fraxinus). I believe this information should have been included in the Resources Planning Act report in order to insure that decision-makers consider these threats in guiding USFS programs.
I also wish the USFS RPA had at least prominently referred readers to Poland et al. Among that study’s key points are:
Invasive (non-native) insects and diseases can reduce productivity of desired species, interactions at other trophic levels, and watershed hydrology. They also impose enormously high management costs.
Some non-native pests potentially threaten the survival of entire tree genera, not just individual species, e.g., emerald ash borer and Dutch elm disease. I add white pine blister rust and laurel wilt.
Emerald ash borer and hemlock woolly adelgid are listed as among the most significant threats to forests in the Eastern US.
White pine blister rust and hemlock woolly adelgid are described as so profoundly affecting ecosystem structure and function as to cause an irreversible change of ecological state.
Restoration of severely impacted forests requires first, controlling the non-native pest, then identifying and enriching – through selection and breeding – levels of genetic resistance in native populations of the impacted host tree. Programs of varying length and success target five-needle pines killed by Cronartium ribicola; Port-Orford cedar killed by the oomycete Phytophthora lateralis; chestnut blight; Dutch elm disease; butternut canker (causal agent Ophiognomonia clavigignenti juglandacearum), emerald ash borer; and hemlock woolly adelgid.
Climate change will almost certainly lead to changes in the distribution of invasive species, as their populations respond to increased variability and longer-term changes in temperature, moisture, and biotic interactions. Predicting how particular species will respond is difficult but essential to developing effective prevention, control, and restoration strategies.
Poland et al. summarizes major bioinvaders in several regions. Each region except Hawai`i (!!) includes tree-killing insects or pathogens.
It is easier to understand the RPA report’s not mentioning priority-setting efforts by two other entities, the Morton Arboretumand International Union for the Conservation of Nature (IUCN). These studies were published in 2021 and their lead entities were not the Forest Service – although the USFS helped to fund the U.S. portion of the studies.
The Morton Arboretum led in the analysis of U.S. tree species. It published studies evaluating the status of tree species belonging to nine genera, considering all threats. The Morton study ranked as of conservation concern one third of native pine species; 31% of native oak species; significant proportion of species in the Lauraceae. The report on American beech — the only North American species in the genus Fagus – made no mention of beech leaf disease – despite it being a major concern in Ohio – only two states away from the location of the Morton Arboretum near Chicago.
valley oak (Quercus lobata) in Alameda Co, California; photo by Belinda Lo via Flickr
Most of the species listed by the Morton Arboretum are of conservation concern because of their small populations and restricted ranges. The report’s coverage of native pests is inconsistent, spotty, and sometimes focuses on odd examples.
Tree Species’ Regeneration
Too late for consideration by the authors of the USFS RPA report come new studies by Potter and Riitters that evaluate species at risk due to poor regeneration. This effort evaluated 280 forest tree species native to the continental United States – two-thirds of the species evaluated in the Kevin Potter’s earlier analysis of pest impacts.
The results of Potter and Riitters 2023 only partially matched those of the IUCN/Morton studies. The Morton study did not mention three genera with the highest proportions of poorly reproducing species according to Potter and Riitters: Platanus,Nyssa, and Juniperus. Potter, Morton, and the IUCN largely agree on the proportion of Pinus species at risk. Potter et al. 2023 found about 11% of oak species to be reproducing poorly, while Morton designated a third of 91 oak species to be of conservation concern.
I believe Potter and Riitters and the Morton study agree that the Southeast and California are geographic hot spots of tree species at risk.
Potter and Riiters found that several species with wide distributions might be at risk because they are reproducing at inadequate rates. Three of these exhibit poor reproduction across their full range: Populus deltoids (eastern cottonwood), Platanus occidentalis (American sycamore), and ponderosa pine(Pinus ponderosa). Four more species are reported to exhibit poor reproduction rates in all seed zones in which they grow (the difference from the former group is not explained). These are two Juniperus,Pinus pungens, and Quercus lobata. As I point out in my earlier blog, valley oak is also under attack by the Mediterranean oak borer.
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. Proceedings of the National Academy of Sciences. Vol. 116, No. 35. August 27, 2019.
Lugo, A.E., J.E. Smith, K.M. Potter, H. Marcano Vega, and C.M. Kurtz. 2022. The Contribution of Nonnative Tree Species to the Structure and Composition of Forests in the Conterminous United States in Comparison with Tropical Islands in the Pacific and Caribbean. USDA USFS General Technical Report IITF-54
Poland, T.M., T. Patel-Weynand, D.M. Finch, C.F. Miniat, D.C. Hayes, V.M. Lopez, eds. 2021. Invasive Species in Forests and Rangelands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. Available gratis at https://link.springer.com/book/10.1007/978-3-030-45367-1
Potter, K.M., M.E. Escanferla, R.M. Jetton, G. Man, and B.S. Crane. 2019. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation.
Potter, K.M. and Riitters, K. 2023. A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests 2022, 13, 19. https://doi.org/10.3390/f13010019
United States Department of Agriculture Forest Service. 2023. Future of America’s Forests and Rangelands: The Forest Service 2020 Resource Planning Act Assessment. GTR-WO-102 July 2023
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm