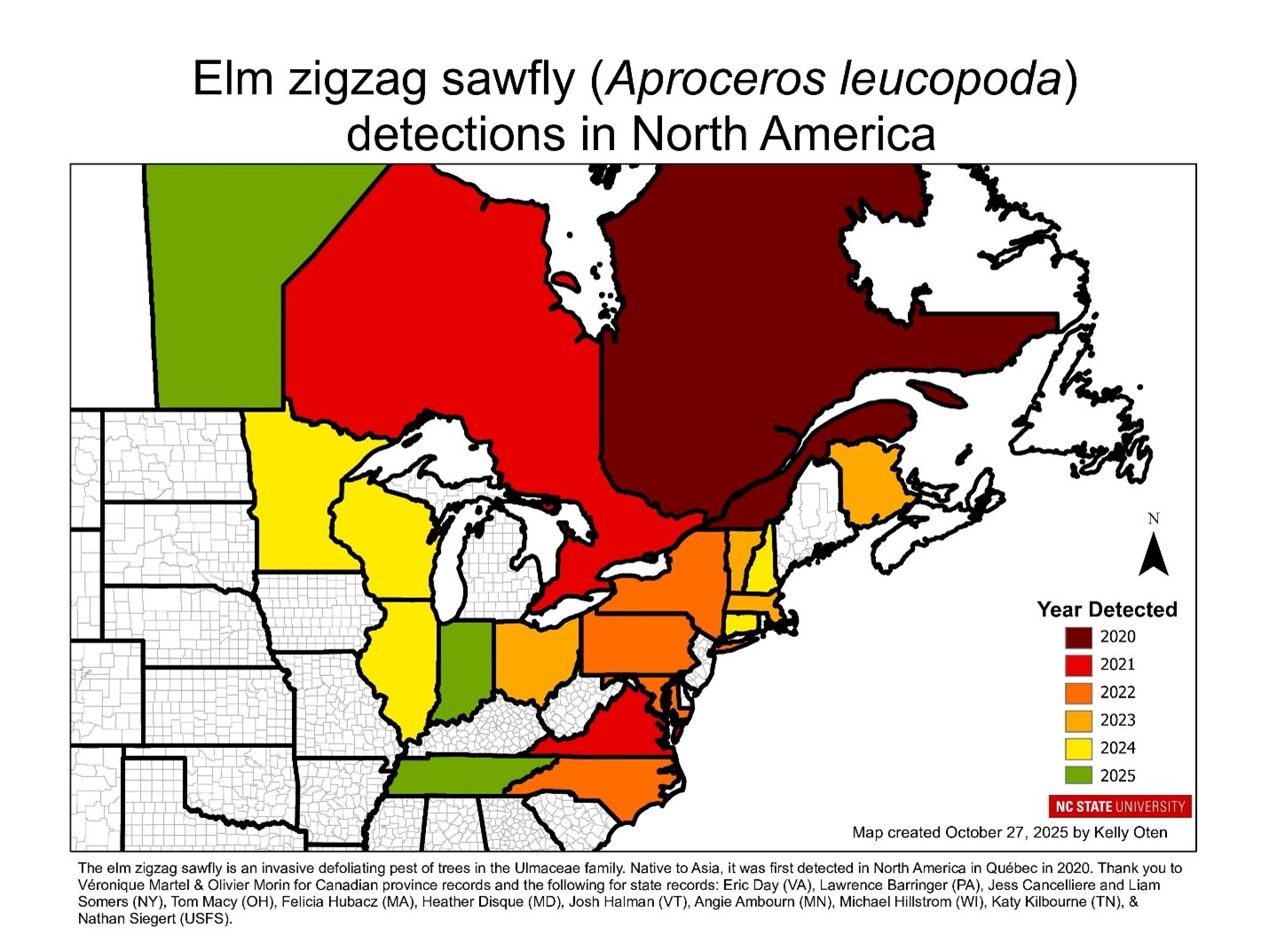
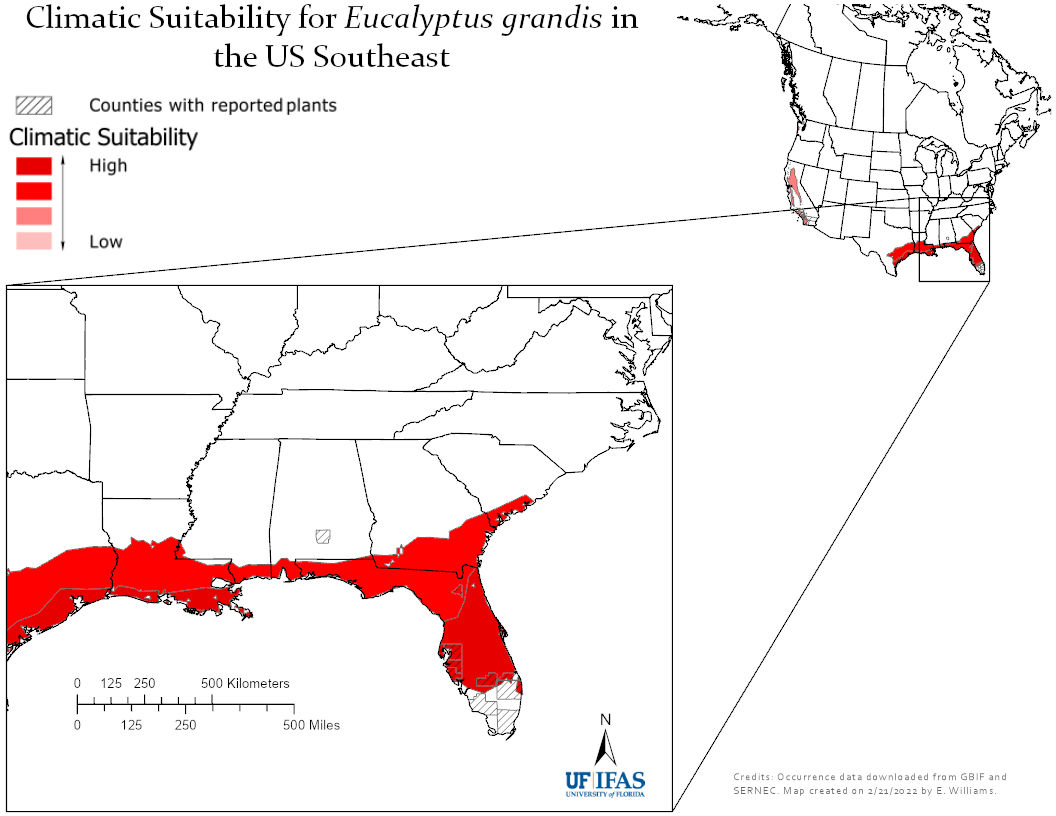
Studies of forest ecosystems in eastern North America that claim to be comprehensive still too often make no reference to invasive species – pests, earthworms, or plants. I try here to bridge these gaps.
Akresh et al. (2023) conducted a meta-analysis of bird species’ use of forests as nesting habitat. They applied the Partners-in-Flight to evaluate the community-wide bird conservation values of unmanaged forests compared to various levels of tree removal by harvest. Because of the decline of many bird species that prefer shrubland or early-successional stands, their process gave highest ranks to management approaches that retained 40%–70% of the canopy trees.
Their study notes that habitats for shrubland birds comprise only about 6% of forests in the eastern U.S. They don’t provide data for southeastern Canada. But hasn’t this scarcity of open upland, non-wetland, habitats in this region been true for thousands of years?
The type of forest that undoubtedly has shrunk significantly in recent centuries is “virgin” (or old-growth or late-seral) forests. As Akresh et al. (2023) report, contemporary closed-canopy forests in eastern North America are predominantly structurally homogeneous, mid-seral, even-aged, stands that have regenerated on land previously cleared for either agriculture or timber. These forests are much younger from a forest developmental perspective than precolonial forests; they lack the latter’s range of tree fall gap sizes and multiple age-classes. The tiny fraction of eastern forests that are in the late-seral stage might have higher species richness and conservation value for birds, but since they are usually not under management, Akresh et al. (2023) did not include that question in their analysis.
Akresh et al. (2023) list the bird species whose density appears to be closely linked to various tree canopy densities. For example, ovenbirds and brown creepers promptly decline in abundance in response to any amount of tree harvesting. Two other species — wood thrush and cerulean warbler — have declined steeply range-wide in recent decades. Nesting densities of three of these four species (excluding the warbler) are significantly higher in areas harvested in ways that retain a greater percentage of trees. Densities of another five bird species (Acadian flycatcher, hermit thrush, black-throated green warbler, and red-breasted nuthatch) are also higher in areas with a greater proportion of trees retained.
Another nine species had a more complex relationship with tree densities but still had lower densities in stands with low tree retention. These were blue-gray gnatcatcher, blue-headed vireo, blackburnian warbler, black-throated blue warbler, eastern wood-pewee, least flycatcher, red-eyed vireo, scarlet tanager, and yellow-bellied sapsucker. They found little relationship between bird density and tree retention for five putative mature-forest species (American redstart, great-crested flycatcher, hooded warbler, veery, and yellow-rumped warbler).

Akresh et al. (2023) claim that silviculture approaches can be used to restore aspects of the structural and compositional conditions found in old-growth forests to second-growth systems, providing a potential pathway for rapidly increasing the conservation value of these areas for bird species. They advocate reducing canopies moderately via variable retention harvests, shelterwood establishment harvests, and irregular shelterwood systems. This strategy can increase understory vegetation density, which they assert can then increase foraging and nesting opportunities for both many mature-forest bird species and many shrubland birds.
I am skeptical; it is much easier to create openings in the canopy than to “create” large trees supporting cavities and associated fauna and flora utilized by some bird species. The authors do advise managers that late-seral, unharvested stands can provide important habitat for old-growth-dependent taxa and any intensive forestry should also take into account other factors.

In addition, often the understory vegetation that responds to the more open environment will be invasive non-native plants. Already about half of eastern U.S. forests have been invaded by non-native plants (Oswalt et al. 2016; Kurtz 2023). Many of these are shrubs: honeysuckles, privets, roses, buckthorn. Management of these plants is difficult – especially when opening the canopy to allow light to reach the forest floor. (at www.nivemnic.us, scroll down to “categories”, click on “invasive plants”.) So the question arises, do the non-native plant species adequately substitute for native shrubs in providing resources needed by those birds?
Maybe. Gleditsch and Carlo (2014) found that a shrub layer dominated by non-native honeysuckle shrubs (Lonicera species) does support nesting populations of several common species, especially catbird (Dumetella carolinensis), American robin (Turdus migratorius ), and northern cardinal (Cardinalis cardinalis). However, they did not consider the species of concern to Akresh et al. (2023) – the rare species that prefer open-canopy, early-successional communities. So they do not inform us whether these high-priority species can utilize shrublands dominated by non-native species. Gleditsch and Carlo (2014) apparently did not find nests of several species considered to be associated with mature forests. So, again, these forests’ value for conservation remains unclear. Gleditsch and Carlo (2014) do counter earlier fears that these non-native shrubs are “traps” for nesting passerine birds. (The concern was that the plants’ structure facilitated nest raiding by predators.) They say, instead, that these plants’ effects are species-specific, context-dependent, and often a mix of both positive and negative outcomes.

Akresh et al. (2023) also do not address the impact of browsing by super-abundant deer. Others (at www.nivemnic.us, scroll down to “categories”, click on “deer”.) have demonstrated that interactions of deer predation with invasive plants is especially damaging to native flora. Considering forests from Virginia to Maine, Miller et al. (2023) advise opening the canopy or subcanopy of forests to promote tree regeneration where deer and invasive shrubs overlap only where deer are controlled.
I have seen no recent analyses of the impact of widespread pest-caused tree mortality beyond some early efforts focused on eastern hemlocks and on high-altitude whitebark pines.
SOURCES
Akresh, M.E., D.I. King, S.L. McInvale, J.L. Larkin, and A.W. D’Amato. 2023. “Effects of Forest Management on the Conservation of Bird Communities in E North America: A Meta-Analysis.” Ecosphere 14(1):e4315. https://doi.org/10.1002/ecs2.4315
Gleditsch, J.M. and T.A. Carlo. 2014. Living with Aliens: Effects of Invasive Honeysuckles on Avian Nesting. PLOS One. September 2014. Volume Nine Issue Nine. E107120
Miller, K.M., S.J. Perles, J.P. Schmit, E.R. Matthews, M.R. Marshall. 2023. Overabundant deer and invasive plants drive widespread regeneration debt in eastern United States national parks. Ecological Applications. 2023;33:e2837. https://onlinelibrary.wiley.com/r/eap
Oswalt, C.M., S. Fei, Q. Guo, B.V. Iannone III, S.N. Oswalt, B.C. Pijanowski, K.M. Potter. 2016. A subcontinental view of forest plant invasions. NeoBiota. 24:49-54 http://www.srs.fs.usda.gov/pubs/48489
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Or