Wisteria floribunda; photo by Jack Stane via Wikimedia
For decades, it has been clear that deliberate introduction of plant species for cultivation plays a central role in the early stages of bioinvasion by plants (and associated insects, plant pathogens, earthworms … even vertebrates. Viz. coqui frogs in Hawai`i.)
Repeatedly over the two plus decades since Sarah Reichard demonstrated this role of ornamental horticulture (see Reichard and White 2001 and Mack 2000), new studies have provided corroborative details. Publications during the past two years show the risks we are still accepting in the United States. Will we act to protect our environment?
Fertakos and Bradley (2024) found that species were likely to establish if they were introduced to as few as eight locations. However, introduction history was not a strong predictor of an established species’ ultimate invasive success. They suggest that other characteristics, like plant traits and local-scale processes (e.g., interspecific interactions), may better predict whether a plant becomes invasive.
Kinlock et al. (2025) also found that plant species that were cultivated longer or were sold by more catalogs were more likely to have “naturalized”. This conclusion was based on analysis of the behavior of nearly 4,000 species sold in nursery and seed catalogs in the continental United States over 200 years. Nearly 41% of these species naturalized somewhere in the “lower 48” states. Unfortunately, they do not discuss what proportion of these species are truly damaging invaders.
Neither Fertakos and Bradley (2024) nor Kinlock et al. (2025) mention the concept of a lag between a species’ establishment and recognized symptoms of invasiveness. Has this concept been repudiated?
Evans et al. (2024) were focused on analyzing which regions of the eastern United States are likely to suffer the worst plant invasions under climate change. In this context, they worry that people will assist non-native plant species’ movement to newly suitable habitats. Evans et al. urge prioritizing for state regulation species in the ornamental trade that are projected to remain or become abundant under the new climate conditions. They say we Americans are poorly prepared to take this action, however, because plant sales are so poorly regulated and only 10% of land managers in eastern North America monitor for new invasive taxa. They say this is because the managing agencies lack of funding and personnel. After 2025’s losses of programs, appropriations, grants, and staff, this deficit is probably worse – not just for federal agencies but also the many state, local, and volunteer programs that have been supported by federal funding.
Beaury et al. (2024) investigated whether plant species recognized as invasive are sold in the same locations as where they are invasive. They found that half of the 89 species named as invasive were sold by a nursery within 21km of an observed record of invasion. The authors say that data gaps mean that these findings underestimate the number of species sold near locations of documented invasions. They warn that at least 25 species are sold by one or more nurseries located in an area that is currently unsuitable for those species, but that will become more suitable for invasion as temperatures warm. Like Evans et al. (2024), they urge proactive regulation to limit these species’ spread.
burning bush Euonymus, Japanese honeysuckle, & English ivy invading a bottomland hardwood site in Fairfax County, Virgina; photo by F.T. Campbell
U.S. Regulatory response is completely inadequate
Beaury et al. (2023) call for regulating the nursery trade in a manner consistent with the scope of the horticultural trade – sales by both e-commerce and brick and mortar stores go to customers far outside a specific state’s jurisdiction. Despite the interstate nature of the trade, sales of horticultural plants are regulated primarily by state governments. Even when a state does restrict the sale of a specified list of invasive plants, the regulations are outdated, tend to include only a few weeds that plague agriculture rather than those that invade natural systems, or are irregularly enforced. The result is a checkerboard of places where a species may legally be offered for sale next to places where that sale is prohibited. Finally, the regulations are reactive; they rarely include plants in anticipation of their spread to new areas. Beaury et al. (2023) call this as a missed opportunity to reduce the likelihood of ornamental escapes.
Evans et al. (2024) also note that online plant sales are relatively unregulated, and state regulations are inconsistent.
Under the Constitution, the appropriate entity for regulating interstate commerce is the federal government. The U.S. Department of Agriculture’s Animal and Plant Health Inspection Service is responsible for populating and managing the federal noxious weed list. Unfortunately, APHIS lists only those taxa that qualify as quarantine pests under the definition of that term in the International Plant Protection Convention (IPPC) Glossary of Phytosanitary Terms. This means that the taxon is either not yet present in the United States or, if present, is not widely distributed and is being officially controlled. Under these criteria, the federal noxious weed list is required to exclude nearly all the invasive plant species sold by the nursery trade.
To counter this enormous regulatory failure, many associations – native plant societies, regional or state invasive plant councils, etc. – publish their own lists of invasive plants. They often encourage their members and the public to either avoid planting these species voluntarily or to plant predominantly native plants. Also, these stakeholders urge nurseries to halt sales of invasive species voluntarily. Dr. Douglas Tallamy points out that even non-invasive, non-native plants disrupt food webs.
These voluntary efforts have yielded some success. But they have not resulted in adequate protection for our ecosystems.
Will Americans choose to invigorate the regulatory system? At a minimum, can we urge neighboring states to adopt a regional approach? More difficult, but also more effective, would be to persuade Congress to strengthen APHIS’ invasive plant regulations to outlaw interstate sales of at least those species documented to be invasive.
Cortadera selloana; picture by Alex Borland via PublicDomainPictures.net
Do you have other suggestions?
SOURCES
Beaury, E.M., J.M. Allen, A.E. Evans, M.E. Fertakos, W.G. Pfadenhauer, B.A. Bradley. 2023. Horticulture could facilitate invasive plant range infilling and range expansion with climate change. BioScience 2023 0 1-8 https://doi.org/10.1093/biosci/biad069
Evans, A.E., C.S. Jarnevich, E.M. Beaury, P.S. Engelstad, N.B. Teich, J.M. LaRoe, B.A. Bradley. 2024. Shifting hotspots: Climate change projected to drive contractions and expansions of invasive plant abundance habitats. Diversity and Distributions 2024;30:4154
Fertakos, M.E. and B.A. Bradley. 2024. Propagule pressure from historic U.S. plant sales explains establishment but not invasion. Public? doi: 10.1111/ele.14494.
Fridley, J.D., P.J. Bellingham, D. Closset-Kopp, C.C. Daehler, M.S. Dechoum, P.H. Martin, H.T. Murphy, J. Rojas- Sandoval, D. Tng. 2025. A general hypothesis of forest invasions by woody plants based on whole-plant carbon economics.
Kinlock, N.L., D.W. Adams, W. Dawson, F. Essl, J. Kartesz, H. Kreft, M. Nishino, Jan Pergl, P. Pyšek, P. Weigelt and M. van Kleunen. 2025. Naturalization of ornamental plants in the United States depends on cultivation and historical land cover context. Ecography 2025: e07748 doi: 10.1002/ecog.07748
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
elm zigzag sawfly larvae feeding on an elm leaf; photo by Delaney Serpan
As one of the newest – and most unique – invasive insects, elm zigzag sawfly (EZS; Aproceros leucopoda) has been making headlines across the eastern U.S. and Canada since 2020. The defoliating pest was first confirmed in North America in Québec, Canada and has since spread rapidly across many states and provinces. As its name suggests, EZS larvae feed primarily on elm in a distinctive zigzag pattern. Moving inwards from the leaf edge, the larvae can eventually consume nearly the entire leaf, leaving nothing but the midrib and a few lateral veins behind. Defoliation from EZS can range from nearly undetectable to 100% canopy defoliation of a mature tree.
EZS is a multivoltine insect, meaning it can have multiple generations in a single growing season. In Europe, where EZS has been invasive since 2003, 1 to 4 generations are common though up to 6 generations have been recorded. In the U.S., many regions document up to 5 generations per year.
In the early spring, EZS emerges from the soil where it has overwintered. They reproduce parthenogenetically- a form of asexual reproduction- allowing them to lay eggs immediately following adult emergence. Each individual is able to lay up to 49 eggs, drastically increasing EZS reproductive potential. Once the eggs hatch, the larvae begin feeding on the foliage until they are ready to pupate. At that point, the larva may build a summer cocoon, attached to a nearby object such as a branch or fence post. Four to 7 days later, an adult will emerge. The entire life cycle only takes 3 to 6 weeks. Alternatively, the larva could drop to the soil beneath the tree’s canopy where it will build its winter cocoon and overwinter, waiting to repeat the cycle the following spring. A small portion of each generation create overwintering cocoons.
EZS summer cocoons attached to the underside of a leaf with evidence of larval feeding; photo by Delaney Serpan
Where is EZS now?
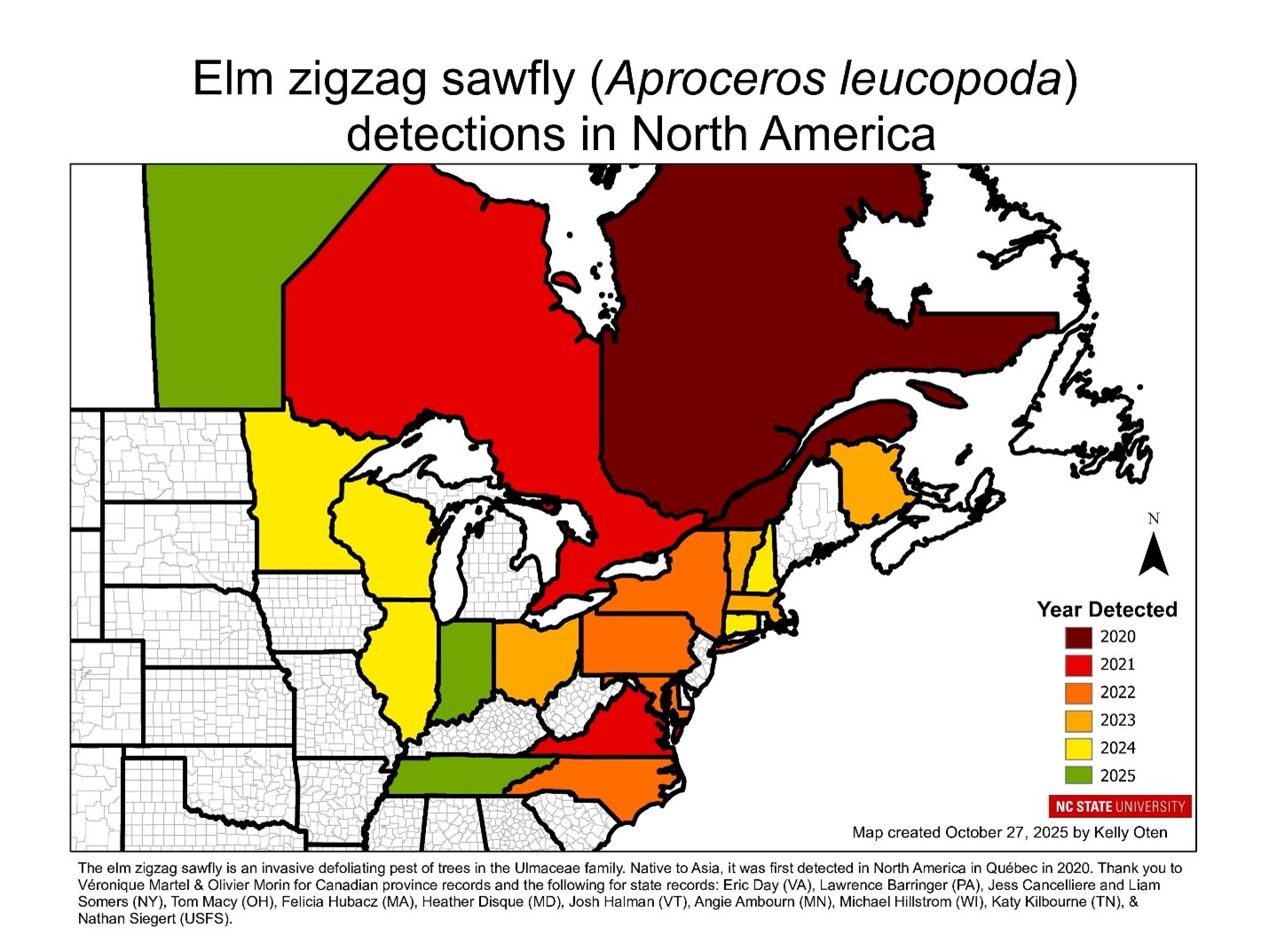
As of the end of 2025, EZS can be found in 15 states and 4 provinces as far west as Minnesota and Manitoba and as far south as North Carolina and Tennessee.
map of states/provinces with official EZS detections;
Invasion pathways in North America are currently unknown; however, EZS has been documented attaching its summer cocoons to truck wheel wells and other objects which may be moved. The subsequent movement of these objects can potentially contribute to EZS spread. It has also been suggested that infested elm nursery stock or potted soil of any plants could be a potential pathway for EZS, but more research is needed to fully understand this.
EZS cocoons on truck – under side mirror & wheel well; photos by Jared Beach, adapted from Oten et al. 2025
How does EZS affect the trees?
Defoliating pests typically decrease the aesthetic value of trees but leave the host largely unharmed. Across Europe and its native range of eastern Asia, EZS defoliation is relatively minimal, with the occasional severe outbreak resulting in total defoliation of a tree. Resulting branch dieback is even more uncommon.
When EZS was first found in North America, particularly North Carolina and Virginia, there were initial concerns about the implications of a warmer climate accelerating development. Like most insects, EZS development is related to temperature; a warmer climate allows for faster insect development. It was hypothesized that a longer growing season could allow for faster population growth and potentially more damage to host trees. At this point, it is still unclear if this will consistently occur in the southern extent of the range. In North Carolina, reported damage has varied widely since it was found there in 2023. Some trees have been 75% defoliated or more multiple years in a row and are exhibiting upwards of 20% branch dieback after just 3 years. However, trees with less than 10% defoliation and no branch dieback have also been recorded.
Since its first detection in North America, researchers have been working to better understand how this pest will affect stakeholders. They’ve been conducting research on the phenology and voltinism of EZS, exploring novel host associations, and evaluating management techniques. Here’s what they’ve learned so far.
A severely defoliated American elm in Surry County, N.C. Photo by Delaney Serpan
First, the bad news.
Elm zigzag sawfly has recently been found to feed on Japanese zelkova (Zelkova serrata), another common ornamental planting within the Ulmaceae family. However, it is important to note that Japanese zelkova is likely not a preferred host. It is suggested that while EZS can complete its life cycle on Japanese zelkova, it will do so only when no other suitable host is present. Researchers are continuing to explore this novel host association.
But help is on the way! There are management recommendations to control elm zigzag sawfly.
Research conducted at North Carolina State University has determined that soil injections of imidacloprid or dinotefuran at label rate are effective methods to significantly reduce larval populations on infested trees. Both active ingredients are easily accessible to landowners and can provide at least one year of protection against EZS. There is ongoing research to explore more treatment options, including augmentative biocontrol.
What can you do about elm zigzag sawfly?
If you are in an EZS-infested region, check vehicles or outdoor items before moving them.
And if you find EZS, report it! To best manage and prevent the spread of EZS, forest health professionals need to know where it is. Elm zigzag sawfly is the only insect that feeds in the unique zigzag pattern on elm trees. If you see the diagnostic feeding pattern, take a picture of it and contact your county’s local Extension agent or state forestry agency to report it.
Invited blog posted by Delaney Serpan
Delaney Serpan is a second-year Ph.D. student in the Forest Health Lab at NC State University, where she studies elm zigzag sawfly biology and management. She first began working with elm zigzag sawfly as an undergraduate researcher shortly after it was detected in North Carolina for the first time. Working with a novel invasive species on the leading edge of its invasion has been incredibly rewarding. Her work aims to provide accessible management techniques to stakeholders while also protecting elms, an already imperiled species, from further damage.
CISP welcomes comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
It used to be thought that closed-canopy forests are resistant to bioinvasion because of the low light availability and relatively infrequent disturbance. Yet many are badly invaded! (On this site, scroll down past the Archives, choose “invasive plants” category.)
Nor is it just temperate forests in North America. Subtropical and tropical forests have also been invaded, as have the temperate forests of South America and, to a lesser extent, Europe. Temperate forests in Asia are less invaded; boreal forests very little (Fridley et al. 2025; see full citation at the end of this blog).
Fridley et al. (2025) have proposed a conceptual model to explain how this happens: “superinvaders” – a special class of woody plants – that achieve competitive dominance across a wide range of forest conditions. The “superinvaders” pose especially grave threats to native biodiversity because they use life-history strategies unlike those of early successional native species.
The result is that existing invasion and succession theories poorly predict which forests are most invasible and by which species. This failure undermines pest risk analyses and early detection.
Fridley et al. have raised lots of interesting ideas – some of which cannot yet be demonstrated by observations.
Temperate forests of North and South America are increasingly dominated by non-native deciduous and semi-evergreen shrubs and trees that combine fast growth rate in high light and high survivorship in forest interiors. These traits enable them to outcompete the native species. Many invaders also produce many more seeds than co-occurring native species. Similar traits are found in the successful non-native plants in subtropical and tropical forests.
Amur honeysuckle via Flickr
Fridley et al. stress that shade tolerance alone does not endow the invasive plants with sufficiently large advantages in their competition with native species. The forest “superinvader” phenotype must combine this ability to persist in shade with high maximum growth rate and high fecundity when conditions become favorable for reproduction.
They explain the invader’s competitive advantage as the result of their experiencing relatively fewer carbon costs because of enemy release in the novel environment, recent environmental changes that alleviate some stress formerly present in the novel environment, or phylogenetic constraints on the local flora that limit natives’ resource-use efficiency. The non-native plant species enjoy this advantage regardless of whether they also possess other competitive mechanisms, e.g., production of allelopathic compounds, denser growth or shading, greater apparent quantum yield. However, Fridley et al. concede that they lack sufficient evidence to incorporate these other competitive mechanisms into their model.
Since any reduction in carbon costs will enhance both shade tolerance and growth rate when light levels are high, these “superinvaders” can outcompete native species in either situation.
To support these concepts, Fridley et al. note that increased abundance of invaders following disturbance is more pronounced in forests than other habitats. They suggest this is because of the much greater magnitude of change in light levels in forests than in open habitats such as grasslands.
They propose that an analogous situation applies to the presence or absence of mutualist microbial associations, although existing studies are insufficient to reach conclusions about the role of carbon allocation to mycorrhizae in the “superinvader” phenotype. The extent to which these forest invasions alter ecosystem-level carbon dynamics, especially soil processes and litter decomposition is also largely unknown.
Fridley et al. emphasize the role of carbon costs in driving both growth rate and whole-plant light compensation point. This point is defined as the light level at which carbon gain through photosynthesis balances carbon losses from tissue respiration (maintenance and growth) and turnover (shedding and loss from disturbance and herbivory).
To survive in low-light conditions, plants must minimize tissue respiration and turnover. The traits that enable those behaviors have been thought to prevent rapid growth and competitive dominance in high-light conditions. However, the “superinvaders” defy this trade-off by growing faster than most co-occurring native species when light is abundant. Fridley et al. say this is because the plants’ reduced carbon costs enhance competitive advantage in both shade and adequate light conditions.
Fridley et al. name several reasons why a native plant’s carbon costs might exceed those of an introduced species. First on the list is either herbivory or investment in defensive traits. Native plants might be challenged by rising abundance or consumption rates of native or introduced herbivores, such as deer or seed predators, that avoid the introduced species.
A second factor is that the non-native species expends fewer resources to sustain adaptations that confer resistance to other stresses, such as drought or freezing. If a long-standing stress is weakened by global change processes (e.g., atmospheric CO2 levels, growing season duration, precipitation levels and seasonality, suppression of fire, atmospheric nitrogen deposition), a non-native plant that lacks defenses against that now-weakened stress will have a lower carbon cost and therefore an advantage. In some cases, the non-native species might benefit directly from these changes, e.g., droughts.
In some regions phylogenetic constraints have limited evolution of adaptive solutions to various biotic and abiotic stresses. This is most obvious on tropical oceanic islands. Fridley et al. report that native trees in Hawaiian montane rain forests are less energy-efficient conducting photosynthesis than are the invaders. However, this phenomenon also occurs on continents. Two continents’ floras might experience different climatic histories even when at they are at similar latitudes. For example, Eurasian woody species leaf out earlier and senesce later than North American trees and shrubs – possibly as a result of more predictable spring and autumnal climate across Eurasia. They name as one example Norwegian maple (Acer platanoides) in North America.
The future is uncertain
Fridley et al. consider enemy release to be a key factor in these shrub invasions of closed-canopy forests. Therefore, if enemy release decays over time because the introduced plant species accumulate pests, or the forest environment shifts to favor more stress-tolerant phenotypes of some native species, then the dominance of superinvaders will decline. If, on the other hand, resource enrichment continues, e.g., nitrogen deposition and elevated CO2, the impacts of woody invaders – present or newly introduced – might continue to rise. The likelihood that additional introductions of more resource-efficient species will continue to damage floras of oceanic islands.
Implications for risk assessments and management
Fridley et al. warn that habitat-matching criteria might be unreliable predictors of forest invasiveness. Among several examples of species that are invasive in interior forest systems in a novel region that do not exhibit this trait in their native range is red oak (Quercus rubra). It is locally dominant in both natural and managed forests in central Europe while in North America, red oak struggles to regenerate in closed-canopy forests. They suggest that Q. rubra in Europe has escaped seedling pathogens present in its native range in North America.
red oak sapling in swampy forest in Virginia; photo by F.T. Campbell
Fridley et al. call for research on traits they have identified as important but that are rarely measured in invasion studies. These include rates of tissue loss and respiratory processes above- and below- ground, plant carbon allocation to tissues and processes, and the whole-plant light compensation points of native and invasive plant species.
The Fridley et al. hypothesis has been supported explicitly by Kinlock et al. (2025). This article says that consistent findings have been reported by earlier small-scale studies in U.S. forests.
I ask for your input on how well the Fridley et al. hypothesis explains shrub and tree invasions in American forests – including those on tropical islands! Is it helpful? Is APHIS incorporating these ideas into plant risk assessments? –
Fridley et al. take pains to reiterate the long-accepted importance of ornamental horticulture in explaining invasive plants’ entry and establishment. They do so in the context of concurring that ruderal traits are not universally advantageous; traits’ benefits depend on the landscape into which the species was being introduced.
SOURCES
Fridley, J.D., P.J. Bellingham, D. Closset-Kopp, C.C. Daehler, M.S. Dechoum, P.H. Martin, H.T. Murphy, J. Rojas- Sandoval, D. Tng. 2025. A general hypothesis of forest invasions by woody plants based on whole-plant carbon economics.
Kinlock, N.L., D.W. Adams, W. Dawson, F. Essl, J. Kartesz, H. Kreft, M. Nishino, Jan Pergl, P. Pyšek, P. Weigelt and M. van Kleunen. 2025. Naturalization of ornamental plants in the United States depends on cultivation and historical land cover context. Ecography 2025: e07748 doi: 10.1002/ecog.07748
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Leucanthemum vulgare (ox-eye daisy); ranked by EICAT as “major impact”; photo via picryl
Because urban centers are “hotspots” of species introductions and reservoirs supporting their spread into areas less altered by human activity, a global group of scientists (Richardson et al. 2025) sought to determine whether the same plant species naturalize in urban areas around the world and – if so – where most of those plant species originate.
They chose to pursue this question because urban areas share many interacting environmental and biotic features that they thought might partially overcome the distinct biomes of the continents. These shared features include the prominence of impervious surfaces; increased habitat heterogeneity; eutrophication; fragmentation of any remaining semi-natural habitats; complex human-influenced disturbance regimes; diverse opportunities for dispersal; novel biotic assemblages and interactions; and human facilitation of non-native species’ colonization and local species’ extinction. In addition to the similarities of the receiving ecosystems, these commonalities are facilitated by shared introduction pathways – although Richardson et al. to not pursue this aspect.
The scientists consulted global invasive plant databases to compile a list of 7,792 plant species recorded as naturalized in one or more of 553 urban centers on all six continents (all except Antarctica). Just over 300 species (4%) were reported on all six continents. They call them the “omnipresent” taxa. Further refinement resulted in a list of 96 species that are particularly widespread, defined as being present in more than half of the urban centers of Oceania, North and South America, and Europe. These 96 species are present in a lower proportion of cities in Asia and Africa. Richardson et al. proposed that these species be folded into a new ecological category, the “urban florome”.
I wonder whether this set of species tells us more about biases in the data than the actual “urban florome”. First, 87% of the 96 “most widespread” species (n= 84) are annual or perennial herbs. Only seven tree, six vine, and six shrub or subshrub species were included among the 96 species. In other words, global lists of invasive species are heavily slanted toward species that thrive in disturbance. Is this surprising? As another study (Kinlock et al. 2025) notes, disturbance is ubiquitous!
Second, only a third of the “urban florome” species have been formally evaluated using the Environmental Impact Classification for Alien Taxa (EICAT) system. Of these 32 species, only six were categorized as having a “major” or “massive” impact. Richardson et al. (2025) conclude that many of the species on the most widespread list are human commensals that have few or negligible known impacts.
Still, this finding might underestimate their impacts. First, as noted, two-thirds have not been evaluated. Second, impacts important in urban systems might not be those that increase a species’ rank based on impacts to natural systems (Richardson et al.). Those with substantial nuisance value in the urban setting still require management. Of course, some of the species have severe impacts in both natural and urban ecosystems. For example, Ailanthus altissimacauses major infrastructural damage and pollen allergies, while Robinia pseudoacaciaalters soil fertility. Both reduce species richness.
Ailanthus
I note that these examples are both trees – which constitute only 7 of the 96 species. Fridley et al. report that trees and shrubs have severe impacts in closed forest systems. I suggest that since many of the urban areas in temperate, subtropical, and tropical regions are probably located in formerly forested areas, remnant (semi-)natural stands and even recreational parks have probably been invaded by these high-impact species. Surely that is more important – at least as regards the level/intensity of the non-native plant species’ impact on biodiversity – than the annual weeds growing along highway verges.
Richardson et al. fear that many cities also have substantial invasion debt. The note specifically that due to the heat island effect, species that can now survive only in cities are likely to spread into surrounding rural and natural areas as temps increase. Thus, these species amplify the urban source effect of plant invasions.
Generalities
Richardson et al. call attention to certain parts of the world acting as ‘factories’ for the evolution of plant species that are well equipped to become invasive when intro to new regions. They name Australian woody flora — although only one species, Melia azedarach, is included among the 96 most widespread species. They also name African grasses and Europe (no taxa specified).
Richardson et al. say that while non-native species in urban areas have usually been described as “passengers” taking advantage of anthropomorphic environmental change, bioinvasions are increasingly recognized as drivers of secondary changes that alter the capacity of these ecosystems to deliver key ecosystem services, or even create disservices. These modifications occur in urban as well as more natural environments.
Regional Differences
Richardson et al. developed lists of the most widespread naturalized urban species for each continent (‘continental lists’). Eighty-seven percent of the 96 “most widespread” species are present in cities of North America, 80% in cities of Oceania, and 34% in European cities. Only 17% of the “widespread” species are present in cities of South America, 13% in cities of Africa or Asia.
While there is considerable overlap regarding species found on several continents, Europe’s urban florome differed significantly from those of the other continents.
The principal source region for these naturalizing species was temperate Asia (145 records); followed by Europe (128 records) and Africa (121 records). Lower numbers came from tropical Asia (95 records); South America (54) records; North America (53 records); and Oceania (8 records). Europe has received 50% of its widespread urban invasive species equally from temperate Asia and North America. Africa has received 75% of its widespread urban species from the two Americas equally.
According to these data, Oceania has been a significant contributor only to South America. I am surprised given the publicized problems caused by Australian Acacia and Hakea in South Africa. I guess these trees are more invasive in the vicinity of urban areas rather than in the cities themselves.
Richardson et al. note a highly skewed relationship between North and South America: while 15.4% of species naturalized in South American cities come from North America, only 2.7% of naturalized species in North American cities are from South America.
Lepidium didymum – brassica from South America introduced widely, including throughout California; photo by Miguel A.C. via Pl@ntnet
Richardson et al. found a distinct division between the “Old” and “New” Worlds (defined by whether the soil was historically cultivated by plough vs. hoe). The latter has more naturalized species (9,905 taxa vs 7,923 taxa), although the “Old World” covers a larger area. Citing di Castri (1989), they suggest that the much longer history of intense human-mediated disturbances in Europe might have allowed its flora to adapt to coexist w/ humans. I wonder, however, whether it is just too difficult to distinguish introductions that occurred millennia ago.
Richardson et al. also found an “echo” from European colonization — strengthened by activities of acclimatization societies. The result is that the continents with longer histories of European colonization, i.e., South and North America and Oceania, have more widespread naturalized plant species than do Africa and Asia.
SOURCES
Fridley, J.D., P.J. Bellingham, D. Closset-Kopp, C.C. Daehler, M.S. Dechoum, P.H. Martin, H.T. Murphy, J. Rojas- Sandoval, D. Tng. 2025. A general hypothesis of forest invasions by woody plants based on whole-plant carbon economics.
Kinlock, N.L., D.W. Adams, W. Dawson, F. Essl, J. Kartesz, H. Kreft, M. Nishino, Jan Pergl, P. Pyšek, P. Weigelt and M. van Kleunen. Naturalization of ornamental plants in the United States depends on cultivation and historical land cover context. Ecography 2025: e07748 doi: 10.1002/ecog.077
Richardson, D.M., L.B. Trotta, M.F.J. Aronson, B. Baiser, M.W. Cadotte, M. Carboni, L. Celesti-Grapow, S. Knapp, I. Kühn, A.C. Lacerda de Matos, Z. Lososová, D. Li, F.A. Montaño-Centellas, L.J. Potgieter, R.D. Zenni, P. Pyšek. 2025. Here, There and Everywhere: Widespread Alien Plants in the World’s Urban Ecosystems. Global Ecology and Biogeography, 2025; 34:e70159 https://doi.org/10.1111/geb.70159
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Kevin Saville reports for the Journal of Commerce that containerized imports in 2025 are expected to be only 25.2 million TEUs, a decrease of 1.4% from 2024.
Declines are particularly large in the final months of 2025 since importers frontloaded their purchases to try to get ahead of the Trump Administration’s new tariffs. Imports for the first half of the year were up 3.6% compared with 2024 at 12.53 million TEUs. Thus, Saville’s sources expect November import levels to be 11.6% lower than in November 2024; December’s to be almost 13% lower.
Analysts expect the steeper decline to continue into the new year. Ben Hackett, of Hackett Associates, expects import volumes in the first four months of 2026 to be 10.3%, 8.5%, 16.8% , and 11% lower than the corresponding months a year earlier. The data source covers the ports of Los Angeles/Long Beach, Oakland, & Seattle & Tacoma on the West Coast; New York/New Jersey, the Port of Virginia, Charleston, Savannah, Port Everglades, Miami & Jacksonville on the East Coast; & Houston on the Gulf Coast. These are not all the maritime ports, but they are the major ones.
Another JOC reporter, Michael Angell, quoted several sources as saying they expect import volumes for all of 2026 to be flat or down 2% from 2025. Illustrating the reversal from past trends, The Port Authority of New York and New Jersey expects total container volumes in 2026 to be 8.5 million TEUs, a decline of about 2% from 2024. Since 2016, NY-NJ port container volumes have grown at an annual average of 4.2%.
As I have blogged before — see here and here — these swings in import volumes threaten to undermine programs intended to prevent introductions of wood-boring insects hitching rides in wood packaging material. While the higher volumes arriving from Asia in the first half of 2025 pose the most obvious risk, falling volumes reduce fee-based funding that support port inspectors. Another factor is the shift to suppliers other than China – primarily countries in Southeast Asia. Two beneficiaries of this shift are Vietnam and – at least initially – India. They have much better records of compliance with ISPM#15-mandated treatments for wood packaging link than does China.
A third JOC source reports that while US and European imports are down, trade volumes in Asia, Africa, the Middle East and Latin America are rising. I expect this growing trade to facilitate new pest introductions, although we will have to wait several years to see any data.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
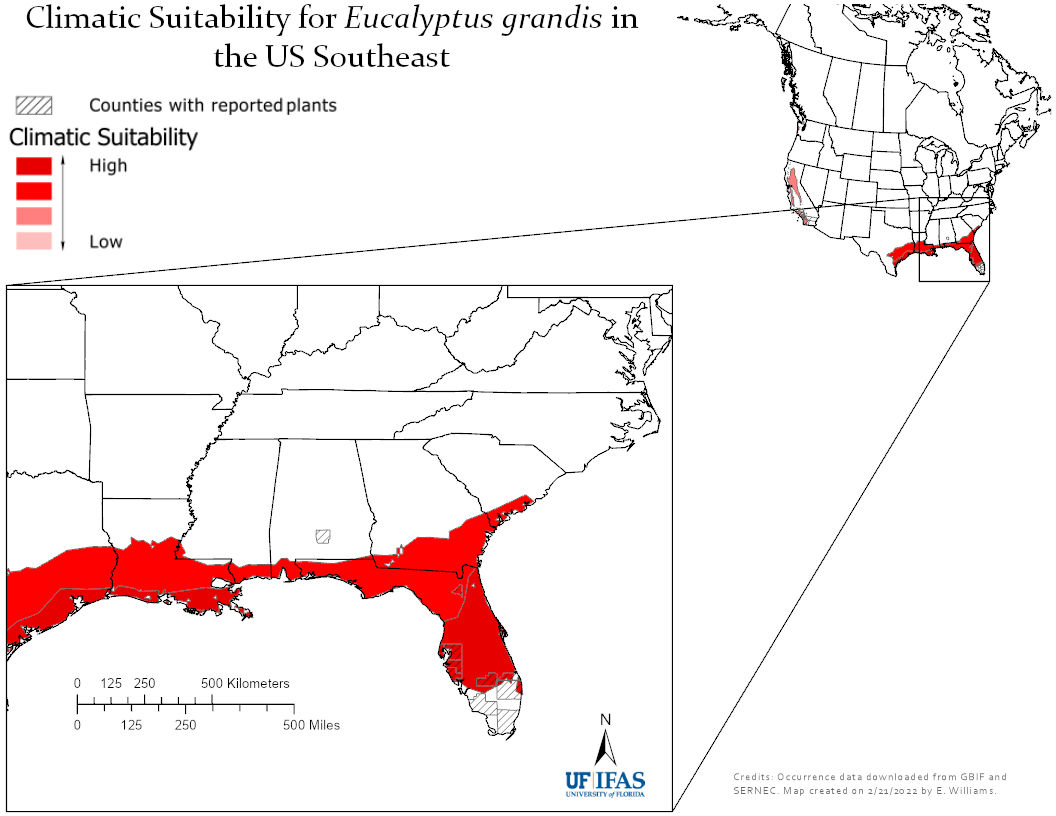
Eucalyptus grandis (in Australia); photo by Poyt448 Peter Woodard via Wikimedia
Deus et al. 2025 (full citation at the end of this blog) have published a review of current knowledge on the invasiveness of trees in the Eucalyptus genus. They report that eucalypt plantations cover more than 30 million ha globally; they could not determine the actual extent more precisely. The area is expanding at an estimated 4% per year. Eucalypts are so popular as timber trees because of their fast growth, ease of management, wood quality and environmental tolerance.
Until recently, trees in the Eucalyptus genus were thought to pose a low invasion risk. This was because these trees have limited seed dispersal, high juvenile mortality, and were expected to lack compatible ectomycorrhizal fungi in novel environments. However, several risk assessments and reports of ongoing invasions in some locations have raised questions. So Deus et al. undertook a literature survey to try to resolve the issue.
One of the risk assessments concerns the United States; see Gordon et al. (2012). This study – completed a dozen years before Deus et al. undertook their literature survey – cited several other sources documenting harmful invasiveness of nearly a dozen species, including Eucalyptus globulus, E. camaldulensis, E. grandis, and E. tereticornis.
Deus et al. found that the limitations listed above actually can be overcome, so they do not prevent invasions:
seeds can disperse farther than 100 meters from parent plants;
high recruitment densities can compensate for the high juvenile mortality; and
ectomychorrhizal fungi can be found in the root systems of introduced eucalypt plants.
In fact, several Eucalyptus species meet criteria defining invasiveness in the Australian Weed Risk Assessment system. Still, Deus et al. found that existing studies cover too few plantations and species to allow an in-depth comprehensive understanding of eucalypts’ invasion ecology.
One reason that eucalypt trees’ invasiveness remains unresolved is that the countries which have established most large Eucalyptus plantations (Brazil, India and China) have not conducted many studies. Instead, most studies have been done in Iberia and South Africa, which together host less than six percent by area of the world’s estate of eucalypt plantations.
Deus et al. say that several possible reasons have been proposed to explain why Eucalyptus is considered to pose an invasion risk by scientists in Iberia and South Africa, but not in Brazil.
The few studies in Brazil were conducted in intensively managed plantations with very short rotations, which are probably less prone to invasion than plantations managed at low intensity levels.
The Brazilian plantations were established 40 to 50 years ago, whereas those in Iberia were introduced ~ 200 years ago.
Iberia experiences recurrent forest fires.
In Brazil, leaf eating ants attack the trees; this might reduce trees’ vigor.
In Brazil, native forests dominate the environs.
Deus et al. say that these hypotheses have never been tested.
Since studies have been conducted in only a few countries, they have evaluated only a few of the species used in plantations. At least 372eucalypt species have been introduced outside their native range; nine species are planted widely. Yet most of the studies reviewed by Deus et al. covered just two species, Eucalyptus globulus (46% of the studies), and E. camaldulensis (33% of the studies). Still, these two widely cultivated species received the highest invasiveness ranking of all species reviewed (65 and 72, respectively). According to Deus et al., these scores are higher than the average score for 32 species of Acacia – a genus considered to be one of the most invasive tree genera in the world.
Other, potentially invasive species, have not received adequate attention. Deus et al. note that E. tereticornis, which is widely planted in China, India and other regions of Southern Asia, has an invasiveness score of 66, placing it second highest in the evaluation. However, only 12 of 140 articles analyzed by Deus et al. addressed this species.
These eucalypts’ high scores result from their potential to hybridize, to naturalize outside their natural habitat, and from high flammability. Other contributing factors are high seed production and ability to resprout after cutting or fire.
The analysis determined that the major drivers for Eucalyptus invasions are soil disturbance, availability of moisture (essential for seedling establishment), and fire. Recruitment density increases with harvesting and tree age; it decreases when the understory is managed. This partially explains why the abandonment of plantations might promote invasions by eucalypts.
Deus et al. fear that there might be a large “invasion debt” in the regions where few studies have been conducted. Assessments for California and Iberian Peninsula indicate that the best areas for cultivation – under either current conditions or expected new environments linked to climate change – are also those most prone to invasion. A further complication is that in some regions it might be difficult to distinguish plants escaping from small plantations from the plantations themselves. They suggest ways to overcome this difficulty: 1) surveys of recruitment along roadside, where trees would not have been planted; 2) genetic analysis of seedlings and possible parents
Another weakness is that that none of the studies considers changes in fire regime, which probably increases the areas prone to invasion.
Deus et al. think it is unlikely that eucalypt invasions will turn out to be as damaging as those of acacias or pines, but that further invasions involving more species and more regions are very likely.
Deus et al. call for considering eucalypt species’ potential invasiveness when developing strategies for the sustainable management of these plantations, including how to manage those that are no longer economically viable.
Status in the United States
The risk in the United States was evaluated by Gordon et al. in 2012. At the time, there were proposals to plant 5,000 to 10,000 ha/year in the Southeast over the next decade.
Gordon et al. adapted the Australian weed risk assessment system to evaluate 38 Eucalyptus taxa then being tested and cultivated in U.S. for pulp, biofuel, and other purposes. Their analysis concluded that 15 of these taxa posed a low risk; 14 taxa posed a high risk; and 9 taxa could not be ranked without further information. The four taxa cultivated most extensively – E. globulus, E. camaldulensis, E. grandis, and E. tereticornis – all had high risk outcomes, as did several other taxa. Gordon et al. thought that these differences reflected both new data and differences in how the assessors reacted to insufficient data.
Gordon et al. warned that novel genotypes with unknown invasiveness were being propagated in the search for increased cold tolerance. This meant that the taxa they had assessed might not indicate of the actual long-term invasion risks associated from this genus. A major source of uncertainty is the long lag time in appearance of evidence of a tree species’ invasiveness. Only one study (as of 2012) had quantified lag time for introduced tree species; it found an average of 170 years from the time introduction to identification of the taxon as invasive. Propagule pressure also influences the lag time and the probability of invasion.
Since the bulk of expanded cultivation was expected to be in the southeast, Gordon et al. recommended that a regional assessment be conducted to more precisely specify the effects of possible differences in phenology, age at reproductive maturity, seed viability, and cold tolerance.
Gordon et al. suggested several actions to reduce the invasion risk. First, selection and breeding strategies could aim to minimize relevant traits – especially eliminating seed production. Second, plantations could be so managed by avoiding cultivation near waterways, harvesting stems before seeds can mature, and restricting the extent of cultivation of any one taxon. More broadly, a fund could be established to cover control costs; growers would contribute the money.
What has happened in the dozen years since the analysis was published? My Google search led to publications from 2013 and earlier. I hope this indicates that no one has funded major expansions. Dr. Gordon reports that most Eucalyptus pulp is imported. ArborGen continues to breed Eucalyptus in Brazil – as I noted earlier, scientists there are not pursuing studies of possible invasiveness of eucalypts.
Still, the regional risk assessment has not been conducted. Worse, Dr. Gordon reports that the Florida Department of Agriculture and Consumer Services has exempted several species [E. amplifolia, E. benthamii, E. dorrigoensis, E. dunnii, E. grandis, E. gunni, E. nitens, E. smithii, and E. urograndis (E. grandisE. urophylla)] from a requirement that growers obtain Non-Native Species Planting Permits. So if the market does take off, there will be no regulation by the state.
At the end of December 2025, Dr. Gordon received information from Florida Division of Plant Industry that no one has applied for a permit to grow Eucalyptus in the state other than under USDA research auspices. So my worst fears have not (yet) come to pass.
I note that in 2022, Potter, Riitters, & Guo ranked Eucalyptus grandis & E. globulus as potentially highly invasive. Their criterion was that at least 75% of stems detected by USFS Forest Inventory and Analysis (FIA) surveys were saplings or seedlings.
SOURCES
Deus, E., D.M. Richardson, F.X. Catry, F.C. Rego, J. Gaspar, M. Nereu, M. Larcombe, B. Potts, J.S. Silva. 2025. Invasion ecology of eucalypts: a review. Biol. Invasions (2025) 27:239 https://doi.org/10/1007/s10530-025-03695-1
Gordon, D.R.,S.L. Flory,.L. Cooper, and S.K. Morris. 2012. Assessing the Invasion Risk of Eucalyptus in the United States Using the Australian Weed Risk Assessment. International Journal of Forestry Research Volume 2012, Article ID 203768, 7 pages doi:10.1155/2012/203768
Potter K.M., Riitters, K.H. & Guo, Q. 2022. NIS tree regeneration indicates regional & national risks from current invasions. Frontiers in Forests & Global Change
doi: 10.3389/ffgc.2022.966407
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
pine plantation near Buenos Aires; photo by Biologicadero via Wikimedia
I have learned about the introduction of a North American woodwasp, Sirex obesus, in Brazil. Forestry interests in South America are worried that this woodwasp will cause significant damage to the pine plantations occupying 4.6 million hectares on the continent.
In July 2023, experts at the Estação Experimental de Ciências Florestais at ESALQ/USP in Itatinga, São Paulo, Brazil, investigated dead and symptomatic trees of several Pinus species and subspecies. They expected the causal agent to be Sirex noctilio – a woodwasp native to Europe and North Africa that has caused considerable damage to South American pine plantations since the 1980s (Wilcken et al.).
However, the pine species attacked were not typical hosts for S. noctilio (in Brazil, loblolly pine Pinus taeda). Instead, the infected trees were Caribbean pines, i.e., Pinus caribaea hondurensis, P. caribaea bahamensis, P. caribaea caribaea, P. maximinoi, P. tecunumani. The responsible woodwasp was identified as Sirex obesus. This species is native to the southwestern United States and northern and central Mexico (Wilcken et al.). This species is closely related to S. californicus (Wilcken et al.).
A second outbreak was found in November ~ 130 km away (still in São Paulo state). Scientists have not determined whether the two São Paulo outbreaks are related. Dr. Villacide reports (pers. comm.) that the two populations genetics have been compared, but he does not have the results.
A third population has been detected in a second, neighboring, state, Minas Gerais (Wilcken to Lantschner and Villacide).
Dr. Villacide (pers. comm.) reports that Brazilian scientists are trying to delimit the extent of the outbreaks. Public and private scientists in other countries with pine plantations have begun developing responses.
This is the first record of S. obesus outside of North America (Wilckens et al.).
Little is known yet about this woodwasp’s probable impact. It is clear that it can oviposit in a wide range of pines. In its native range, S. obesus has been reported on three host species: Pinus ponderosa, P. teocote (twisted-leaf pine), and P. leiophylla (no common name; native to Chihuahua – mostly in Mexico, and border areas of New Mexico and Arizona]. In Brazil, as noted, it has been recorded on other species as well as the hybrids P. caribaea x P. elliottii and P. caribaea x P. tecunumanii (Wilcken et al.).
So for purposes of their risk assessment, Lantschner and Villacide assumed that S. obesus can affect any of the species commonly planted in the region: P. taeda, P. elliottii, P. ponderosa, P. contorta, P. caribaea, P. oocarpa, P. patula, P. radiata, and P. tecunumanii (Lantschner and Villacide).
The risk assessment predicts suitable climatic conditions for invasion by S. obesus in 48% of the areas where South American pine plantation occur, particularly in montane and high-altitude regions along the Andean corridor and central-eastern Brazil. Incorporating other factors – host distribution, proximity to invaded areas, and volume of wood imports from Brazil – identified the most vulnerable areas as in southern Brazil, northeast Argentina, the Argentine Patagonia, and central Chile (Lantschner and Villacide).
pine plantation in Argentina; photo by Tomas Asurmendi via pexels
Preliminary sampling (Wilcken et al.) indicates the impacts could be severe. Mortality varies by species: in the worst cases average mortality approached 43% on P. caribaea hondurensis but only 11% on loblolly pine (P. taeda). They expect mortality rates to increase. Another 30% of P.c. hondurensis trees are dripping resin, a sign of woodwasp oviposition. If these eggs hatch, those larvae will probably kill the affected trees. Such a result would increase total mortality of P.c. hondurensis from 43% to ~ 73%. For P. taeda, the current mortality rate of 11% could rise to 49% as an additional 38% of trees succumb. Following this logic, these areas could experience complete tree mortality within a few years. Given the extent of pine plantations, and possible mortality rates, even a partial spread of S. obesus could lead to significant econ losses.
As second factor is the number of generations per year; the higher the number, the faster woodwasp populations can increase. Wilckens et al. report that adult emergence in Pinus logs maintained in cages indicates that S. obesus could have two or three generations per year.
S. obesus seems to prefer a different climate than S. noctilio. As noted, S. obesus seems to prefer montane and high-altitude climates. S. noctilio is concentrated in lowland temperate and humid regions (Lantschner and Villacide). The newly introduced species might substantially broaden the geographic area where pine plantations might be at risk – although further research is needed to clarify this point.
S. obesus also appears to be spreading at a rapid rate — ~46 km / year. At this rate, Lantschner and Villacide say it could spread throughout all major pine plantation areas in Brazil in less than years.
Sirex woodwasps kill trees by injecting a symbiotic wood decay fungus and a phytotoxic mucus into the tree when ovipositing. The toxin weakens the tree, allowing the fungus to spread, typically killing the tree in as little as three–four months. In North America S. obesus is associated with Amylostereum chailletti. While this species has not yet been confirmed in Brazil, (Wilckens et al.). Brazilian scientists are exploring whether S. obesus might adopt the fungus already present, Amylostereum areolatum, which is associated with S. noctilio.
Two insect species known to feed on woodwasps have emerged from logs infested with S. obesus: Ibalia leucospoides (Hymenoptera: Ibaliidae) and a species of Schlettererius (Hymenoptera: Stephanidae). While these two predators have not proved to be effective controls of woodwasps by themselves, they might become part of a control program. The parasitic nematode, Deladenus siricidicola (Nematoda: Neotylenchidae) used successfully in several South Hemisphere countries to control S. noctilio has not been found in Brazil (Wilckens et al.).
Scientists don’t know the pathway by which S. obesus entered Brazil. Wilckens believes it was via wood packaging; technicians from the Ministry of Agriculture have found some pallets associated with imports that lacked the ISPM#15 mark (Wilckens et al.).
Both Lantschner and Villacide and Wilcken et al. stress the vulnerability of South American pine plantations to introduction of damaging pests. The plantations are reportedly intensively managed, even-aged, regularly spaced monocultures. These conditions can facilitate invasive species establishment and spread by providing abundant host resources and reduced natural enemy pressure. Lantschner and Villacide cite Michael Wingfield that in plantation forestry, introduction of a single pest species can damage large areas of valuable timber.
mortality caused by Sirex noctilio in a pine plantation in Argentina; photo courtesy of Jose Villacide
The family Siricidae contains more than 120 species distributed across the forests of the Northern Hemisphere. In their native ranges they are typically minor or secondary pests (Wilckens et al.). Woodwasps have demonstrated that they can be transported in international commerce – S. noctilio alone has invaded pine stands (native or exotic) in nine countries in Oceania, Africa, and South and North America. Three other species in the family — Urocerus gigas, Urocerus flavicornis and Tremex fuscicornis – have been detected in South America (Wilckens et al.). If each represents a unique threat, countries with widespread pine plantations should enhance their phytosanitary programs. Exporting parties, e.g., the United States and European Union, should assist in efforts to prevent spread of these wood borers. One major step would be to strengthen regulations governing wood packaging material. [To see my criticisms of shortfalls of the ISPM#15 system, scroll down the list of blogs to “Categories” and click on “wood packaging”.]
Lantschner and Villacide cautionthat their assessment is based on a limited record of S. obesus occurrences in its native range. This range might be restricted by factors other than climate, including geographic barriers or biotic interactions (natural enemy pressure or interspecific competition). If so, the species’ potential invasive range might be larger than the climate-based models predict.
Recommendations for management strategies
I applaud Lantschner and Villacide for proposing immediate steps to improve management of the threat posed by introduction of S. obesus. These recommendations should prioritize enhanced phytosanitary inspections of wood products moving between high-risk regions and other South American countries. They suggest that Brazil adopt bilateral agreements with its major trading partners which would specify protocols for woodwaspdetection and quarantines. [Since many of these countries already have established populations of S. noctilio they probably do not have strong phytosanitary measures targeting wood borers at present.] Lantschner and Villacide advise creation of targeted surveillance programs in southern Brazil, northeastern Argentina, Argentine Patagonia, and central Chile. They should focus on sites near major transportation hubs and border crossings. Less intense surveillance should be instituted in regions they classified as medium risk. Again, the focus should be on major points of entry for imported goods and on plantations located near the Brazilian border. They note that preventing spread of S. obesus into new areas will require not only national efforts but also regionally coordinated monitoring, research, and forest health policies.
Lantschner and Villacide also identify priority areas for future research. These include clarifying S. obesus’shost range, the environmental conditions that enable the woodwasp to establish and persist beyond its native range, dispersal rates, and whether S. obesus exhibits pulse-like pop dynamics[long periods of low density interrupted by sudden outbreaks] seen in S. noctilio.
Dr. Villacide (pers. comm.) reports that Brazilian scientists are trying to delimit the extent of the outbreaks. Public and private scientists in other countries with pine plantations have begun developing responses. Dr. Villacide has posted a video from a recent online seminar sponsored by the Southern Cone Forest Health Group. Go to https://youtu.be/uVU6CpFNhlQ?si=lqXtwJTtz5rKXfL3 or https://sanidadforestalconosur.org/
A wider prespective
Dr. Villacide’s attention to Sirex obesus is part of his broader work on pest issues in South America’s commercial plantations. In another publication (Villacide and Fuetealba 2025; full citation at the end of this blog), he explores how to make these plantations sustainable in the face of rising threats from pests – both introduced and native to the region. Dr. Villacide and Alvaro Fuetealba report that every year 1.2 million hectares of plantations in the Southern Cone are affected by pests. Their vulnerability of will be worsened by the extreme weather events expected under climate change.
These plantations present vast areas of homogeneous stands: ~97% of the Southern Cone planted area consists of exotic tree species – mainly Pinus and Eucalyptus. Typical plantations are high density and managed intensively – including thinning, pruning, and fertilizing – to prompt rapid growth. As Villacide and Fuetealba point out, while these practices maximize wood production efficiency, they also lead to biological homogenization and reduced resilience to pests.
They report that pine plantations are under attack by wood and bark borers that have followed pines to the region, including Sirex noctilio, Orthotomicus erosus, and Cyrtogenius luteus; and now the newly detected Sirex obesus (above). At least two fungal pathogens — Fusarium circinatum and Dothistroma septosporum – have also been introduced. The principal threat to pine plantations from native pests comes from leaf-cutting ants (Atta and Acromyrmex).Eucalyptus plantations are plagued by several insects that have arrived from Australia, including Phoracantha semipunctata, Thaumastocoris peregrinus, and Leptocybe invasa. Pests native to the region that attack Eucalyptus are the Chilean carpenter worm (Chilecomadia valdiviana) and the leaf-cutting ants.
Cordilleran cypress; photo by LBM 1948 via Wikimedia
Threat to native conifer
More worrying to me is that introduced pests have entered native forests. Villacide and Fuetealba report that the aphid Cinara cupressi is attacking the native conifer Austrocedrus chilensis. Cordilleran cypress, also called Chilean or Patagonian cedar, is an endemic, monospecific tree in the Cupressaceae family. In southern Argentina and Chile the species forms pure and mixed stands with southern hemisphere beech (Nothofagus spp.) across ~ 160,000 ha. The profile Cinara cupressi on the Global Invasive Species Database is unclear about how many species are in the species complex and their places of origin.
Cordilleran cypress is also under attack by the oomycete Phytophthora austrocedri, an oomycete of unknown origin. This pathogen is of unknown origin. It is now thought to have been present in Argentina since at least the 1960s. P. austrocedri has also been ntroduced to Europe, western Asia, and North America.
Villacide and Fuetealba advocate several actions to might diversify tree species in the plantations to reduce their vulnerability to pests. They note that this recommendation builds on foundational ecological theory, including the resource concentration and natural enemy hypotheses. Diversity-promoting actions should reach beyond any plantation to the landscape level. Managers should consider connectivity of susceptible stands, the number of nutritionally optimal host trees in the landscape, and the availability and quality of hosts in adjacent stands.
Villacide and Fuetealba say mixed plantations can provide additional ecological and economic benefits, such as enhanced stand-level productivity; production of a wider range of commercial and subsistence products; and greater resistance and resilience to natural disturbances, e.g., extreme weather events.
They warn that designing and implementing mixed plantations must reflect ecological interactions and pest dynamics as well as management. There is need for regionally coordinated experimental plantations where scientist could test how variables such as tree species composition, density and spatial arrangement, and silvicultural practices influence pest dynamics, forest productivity, and ecosystem resilience under local conditions. They suggest incorporating sentinel plantings both early-warning systems and decision-support tools at plot and regional scales. Researchers should evaluate pest-specific responses, productivity trade-offs, long-term forest health outcomes under different scenarios.
Since the plantations extend across a multinational region with few natural barriers and uniform silvicultural practices, as well as high levels of trade, so do the pest problems. Therefore, the response must also be regional – e.g., regional experimental plantations and living laboratories. A collaborative approach linking researchers, forest managers, and policymakers is essential to translate experimental findings into practice and develop adaptive, ecol grounded silvicultural strategies. Long-term ecological trials must be embedded in operational contexts and aligned across countries.
SOURCES
Lantschner, V. and J. Villacide. 2025. Invasion Potential of the Recently Established Woodwasp Sirex obesus. Neotropical Entomology. (2025) 54:117 https://doi.org/10.1007/s13744-025-01347-6
Villacide, J. and A. Fuetealba. 2025. Pests in plantations: Challenging traditional productive paradigms in the Southern Cone of America. Forest Ecology and Management 597 (2025) 123127
Wilcken, C.F., T.A. da Mota, C.H. de Oliveir, V.R. de Carvalho, L.A. Benso, J.A. Gabia, S.R.S. Wilcken, E.L. Furtado, N.M. Schiff, M.B. de Camargo, M.F. Ribeiro. 2025. Sirex obesus (Hymenoptera: Siricidae) as invasive pest in pine plantations in Brazil. Scientific Reports. 2025. 15:22522 https://doi.org/10.1038/541598-025-06418-7
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm