beech leaf disease – Not one of the plant pests that APHIS is regulating! Photo by Jennifer Koch, USFS
APHIS has reminded us that 2022 is the agency’s 50th year. In its press release, APHIS claims several accomplishments over this period:
Eradicating plant pests like European grapevine moth and plum pox from the country, while reducing the impact of others plant diseases, including boll weevil and Mediterranean and Mexican fruit flies;
Eradicating serious animal diseases, including highly pathogenic avian influenza, virulent Newcastle disease, and pseudorabies, from the country’s herds and flocks, while reducing the prevalence of other animal diseases like bovine tuberculosis and brucellosis;
Improving care for laboratory animals, exhibited animals and other animals;
Ensuring genetically engineered plants do not pose a risk to plant health, while keeping up with the ever-changing technology in this field;
Reducing the impact of wildlife damage on agriculture and natural resources; and
Ensuring safe trade of agriculture commodities across the globe
APHIS also launched a new page on its website to share a series of visual timelines of its history and important milestones.
APHIS also states that USDA) has declared April 2022 to be Invasive Plant Pest and Disease Awareness Month (IPPDAM). The link Invasive Plant Pest and Disease Awareness Month connects you to APHIS’ webpage. Secretary Vilsack asks people to be alert. He noted particularly the risk that pests will hitch a ride on untreated firewood, outdoor gear and vehicles, and soil, seeds, homegrown produce, and plants.
The notice urges people to:
Familiarize yourself with the invasive pests that are in your area, and their symptoms. [Faith says – also look for pests not “here” yet – early detection!]
When returning from travel overseas, declare all agricultural items to U.S. Customs and Border Protection so they can ensure your items won’t harm U.S. agriculture or the environment.
Don’t move untreated firewood. Buy local or use certified heat-treated firewood, or responsibly gather it on site where permitted.
Source your plants and seeds responsibly. When ordering online, don’t assume items available from foreign retailers are legal to import into the United States. Learn how to safely and legally order plants and seeds online.
Don’t mail homegrown plants, fruits and vegetables. You may live in an area under quarantine for a harmful invasive plant pest. You could inadvertently mail a pest.
When in doubt, contact your local USDA State Plant Health Director’s office to find out what you need to do before buying seeds or plants online from an international vendor or before mailing your homegrown agricultural goods.
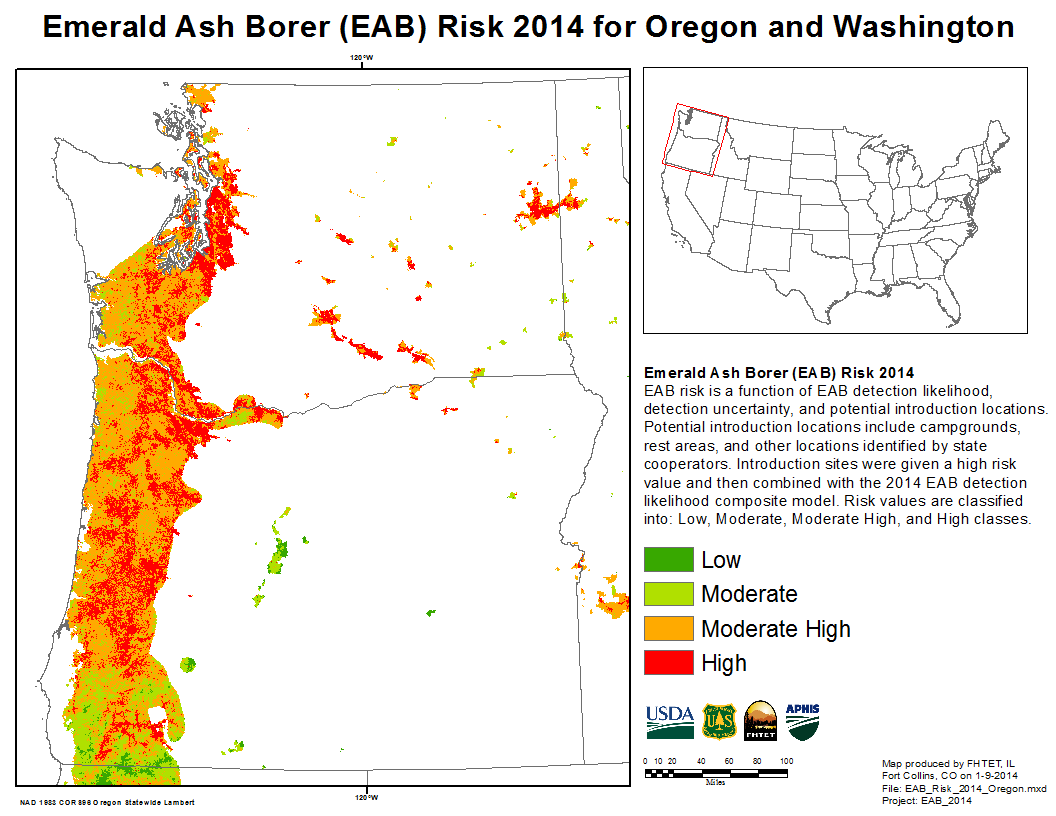
nearly pure stand of Oregon ash in Ankeny National Wildlife Refuge, Oregon; photo by Wyatt Williams, Oregon Department of Forestry
While Michiganders document the impacts of the emerald ash borer (EAB) there, conservationists on the West Coast are jump-starting efforts to save their regional species, Oregon ash (Fraxinus latifolia). Earlier field tests in the Midwest showed that EAB will attack Oregon ash (press release) – something West Coast state would like to counter as early and effectively as possible.
Oregon ash is a wide-ranging species, occurring from California to Washington and possibly into British Columbia. The species has not been studied extensively (it is not a timber species!), but it is clearly an imponearlrtant component of riparian forests. In wetter parts of the Willamette Valley, ash is the predominant tree species. See the photo of the riparian forest in the Ankeny National Wildlife Refuge; this forest is nearly 100% Oregon ash (ODA/ODF EAB Response Plan).
As is true in the Midwest, ash provides important food and habitat resources along creeks and rivers where seasonally high water-tables can exclude nearly all other tree species. Standing and fallen dead ash biomass can alter soil chemistry and affect rates of decomposition, nutrient, and water cycling, i.e., nutrient resource availability for the remaining trees. Gaps in tree canopy can increase soil erosion, stormwater runoff and elevated stream temperatures. In dense stands of Oregon ash, understory vegetation is often sparse, consisting primarily of sedges. The authors of the Response Plan anticipate invasion by non-native plants into canopy gaps caused by the loss of ash trees as a result of an EAB invasion. In Michigan, though, it is the sedges that dominate these gaps.
The Oregon Department of Forestry, the state Department of Agriculture, and other entities have actively participated in “don’t move firewood” campaigns for at least a decade. The Departments of Forestry and Agriculture also led a team that prepared the EAB Response Plan in 2018 (full citation at the end of this blog). It lays out in considerable detail the roles of both government agencies and non-governmental stakeholders. Oregon’s quarantine is broad, covering all insects not on an approved list (Williams, pers. comm.)
California has inspected incoming firewood for years. In April 2021 – after APHIS terminated the federal quarantine on EAB — California Department of Food and Agriculture established a state quarantine on the beetle and articles that could transport it into the state. In doing so, CDFA noted that commercially grown olive trees might also be at risk to EAB.
Washington State operates a statewide trapping program for invasive insects. There has also been significant attention to non-native insect threats to urban forests. These have included a study in 2016 led by the Washington Invasive Species Council (WISC). It involved a partnership of WISC with the Washington Department of Natural Resources Urban and Community Forestry Program as well as and statewide stakeholder meetings [Bush, pers. comm.].
Of these various state-wide initiatives, the institutions in Oregon appear to be most pro-active. The Tualatin Soil and Water Conservation District provided $10,000 to fund some of the genetics work and testing for EAB resistance. Other funding came from the USDA Forest Service Forest Health Protection unit of State and Private Forestry (not from USFS’ Research Program). As described by USFS geneticist Richard Sneizko in an article in the publication TreeLine (full citation at end of blog), participants hope to find at least some level of genetic resistance to EAB. Any such resistance might be deployed in several ways: 1) promoting reproduction by resistant trees to enhance their numbers before EAB gets to Oregon; 2) using seeds from resistant trees for restoration of natural areas; or 3) cross-breeding resistant trees to build genetically diverse stocks of resistant trees for future restoration.
Participants think it is vitally important to work from seeds collected over much of the range of Oregon ash – first, to search for probably very rare resistant trees; and second, to preserve the full diversity of the tree species’ genome so that restored ash will be adapted to the wide variety of conditions in which ash grow.
Participants in this effort include the forest genetics/tree improvement community – specifically, the USDA Forest Service Dorena Genetic Resource Center (located in Cottage Grove, Oregon) and Washington State University at Puyallup Research & Extension Center. Also engaged is the public gardens community, specifically the Huntington Botanical Gardens in San Marino, Los Angeles County. The garden is collecting seed of Oregon and other western ashes from California and Washington State.
The first step in assessing resistance is collecting seed from ash trees across the range of Oregon ash. This began in 2019. Carried out by, inter alia, some USFS and Interior’s Bureau of Land Management units, Oregon State University, citizen scientists [Sniezko] and the Oregon Department of Forestry [press release & Sniezko pers. comm.] Also, some seeds were collected in Washington State in 2020. Additional collections in Oregon are scheduled for 2022.
The collected seeds have been evaluated for vitality and stored by the USFS Dorena Center and at the USFS National Seed Lab (Macon, GA).
Oregon ash planting at Dorena; photo by Emily Boes
The USFS Dorena Center and Washington State University have begun germinating and growing some of the seedlings for various tests of possible resistance. There is concern that the 2021 drought might have killed some of the seedlings in Oregon; those in Washington are not affected. The initial seedlings are mostly from Oregon but there is space to add additional families from a wider geographical area. Experimenters plan to collect data annually on bud break, yearly growth, and any diseases or pests that develop on the trees. (Chastagner pers. comm.)
The next step is systematic testing whether some of the ash show genetic resistance to EAB. Richard Sneizko has sent seedlings of 17 ash families to USFS colleague Dr. Jennifer Koch. She operates a breeding facility in northern Ohio where they can be tested for resistance. Testing is expected to begin this year. [Tree Line]
The Dorena Center is also helping a researcher at Penn State University, Dr. Jill Hamilton, to set up a landscape genomics project. She will evaluate the genetic variability in the species by using leaf samples from about 20 trees from many populations across the Oregon ash’s range (California to British Columbia). This potentially includes a collection from the Dorena population of ash in late Spring 2022. [Sniezko]
These various ash plantings can also be “sentinel” plantings to assist in early detection of newly arriving EAB. [Tree Line]
SOURCES
Bush J. Executive Coordinator | Washington Invasive Species Council
ODF and ODA Emerald Ash Borer Readiness and Response Plan. 2018.
The newsletter is issued by Bonneville Environmental Foundation for a consortium of conservation agencies
Sniezko pers comm Feb 2022 22-2/24
A video explaining the campaign to save Oregon ash is at https://youtu.be/uZmfLrxEA7g or https://youtu.be/S8y-XK285S8
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
results of EAB infestation; photo by Nate Siegert, USFS
A series of studies by Patrick J. Engelken, M. Eric Benbow, Deborah G. McCullough, Nate Siegert, Randall Kolka, Melissa Youngquist and others examine the status of ash (Fraxinus spp.) in the aftermath of the emerald ash borer (EAB) invasion. Initial studies documented the crash of biomass supporting EAB numbers when the large ash trees died (Siegert, Engelken, McCullough. 2021; full citation at end of blog.) More recent studies have focused on bogs and forests in the riparian areas where ash were especially numerous and arguably ecologically most important. I posted a blog about black ash bogs earlier.
I will focus here on the studies in riparian areas of southern and northwest Michigan. Information about impacts in forests of southern Michigan are from Engelken, Benbow and McCullough (2020); information about impacts in northwest Michigan are from Engelken and McCullough (2020). Full citations for both are at the end of the blog.
All study areas had high ash densities before EAB’s arrival. One study (Engelken and McCullough 2020) found ash densities high in the immediate riparian areas (in one case, a strip reaching 100 meters from the streambank) but scattered in surrounding forests.
In all these study areas, populations of mature (reproductive age) ash crashed within 10-15 years after EAB invasion:
In northeast Michigan, EAB carrying capacity was reduced by 94% – 99%; total ash basal area was reduced by 87 – 97.7% (Siegert, Engelken, McCullough. 2021);
In southern Michigan, more than 85% of the basal area of green (F. pennsylvanica) and black ash (F. nigra) had been killed by 2020. An estimated 96% of the overstory ash phloem area had died, thus radically reducing EAB carrying capacity (Engelken, Benbow and McCullough 2020);
In northwest Michigan, more than 95% of the overstory ash have been killed. (Engelken and McCullough 2020).
The worst impact has been on black ash– which plays such an important ecological role in riparian areas and wetlands and has enormous importance in Native American cultures. In all these study areas, there is no stump sprouting by black ash (Siegert, Engelken, McCullough. 2021; Engelken, Benbow and McCullough 2020; Engelken and McCullough 2020). In three watersheds of northwest Michigan where black ash constituted up to a quarter of the overstory species before the EAB invasion, scientists found no black ash recruits, only eight saplings, and a single seedling.
Green ash (F. pennsylvanica) has survived in much higher numbers – so far. However, this species’ ability to grow into reproductive size is still uncertain. In northwest Michigan, green ash saplings are abundant in canopy gaps created by EAB-caused mortality of mature ash. These saplings had established before the EAB invasion so some call them the “orphaned cohort”. However, there are few seedlings of any woody plant species in these gaps because sedges form such dense mats.
Green ash reproduction faces many challenges before persistence of the species can be considered assured. First, populations of EAB – now reduced by the lack of mature ash to support them – might resurge when young ash grow to larger sizes. It is not yet clear the extent to which introduced biocontrol agents and native predators, e.g., woodpeckers, will protect these trees as they grow to reproductive size. Here, again, green ash has an advantage over black ash. While green ash produce seed at a relatively young age, black ash don’t produce seed until they reach 30–40 years. Even then, they produce seeds only sporadically, with intervals of five or more years.
A second challenge is the lack of seed sources – at least until and unless young trees are able to reach reproductive size.
A third challenge is competition for resources from other plants. The canopy gaps eliminate competition for light for the taller plants, i.e., the existing ash saplings. However, the sapling cohort is not supported by a seedling cohort. There are very few seedlings of all woody plant species (including invasive species!). Seed germination is suppressed by the dense mats of wetland-adapted sedges and possibly the higher water tables (which resulted from reduced evapotranspiration following mortality of the mature trees).
Competition for resources is also a factor in the forests outside the immediate riparian zone. There, ash seedlings sprout, but shade created by lateral ingrowth suppresses their growth. In southern Michigan, Engelken, Benbow and McCullough (2020) note that the forests are apparently transitioning from red oak dominated forests to red maple and black cherry dominated forests. This transition is apparently intensified by forest mesophication resulting from reduced fire frequency, decreasing light availability in forest understories and increasing soil moisture content.
Fourth, while stump sprouting of green ash was noted in southern Michigan, in the northwestern forests all the sprouts died. I have already noted the absence of stump sprouting by black ash at all sites.
Beaver & Green Ash in Northern Virginia
photos of beaver feeding on ash saplings in northern Virginia; photos by F.T. Campbell
In spring 2022 I noticed along one stream in northern Virginia that beavers had cut down green ash saplings; McCullough and Siegert report that this does not appear to be a problem in their study areas.
By December 2022, the beaver-cut trees tried to recover: see the sprouts from a stump [below]. (I think deer or rabbits ate the tips of the sprouts.)
The beavers also continued feeding on the ash — the tree photographed in the spring when it was half-chewed through has now been felled and its branches removed [see below].
Ecosystem Impacts, Especially on Streams
Across much of the upper Midwest, massive ash mortality is causing widespread changes in forest systems.
Riparian forests, i.e., areas adjacent to waterways where periodic inundation occurs, are functionally linked to the aquatic systems. Loss of such a significant proportion of the overstory changes the transfer of energy to adjacent waterways that takes the form of inputs of nutrients from leaf litter and coarse woody debris. Intact forests also stabilize stream banks and maintain channel depth by preventing erosion. Forests moderate temperature of the water. Finally, forests with “coarse woody debris” increase habitat structure. These impacts might be especially important along first order streams, (defined as perennial streams that have no permanently flowing tributaries). These streams are too small to buffer the impacts of major tree loss. The scientists say they are uncertain whether these changes continue to affect larger streams downstream.
Unshaded streams have higher water temperatures that can affect populations of fish, in particular salmonids, by delaying migration, reducing egg viability and increasing egg mortality. Higher temperatures can also alter primary productivity of aquatic algae, potentially increasing eutrophication (Engelken and McCullough 2020).
The scientists expect increasing abundance of coarse woody debris in the forests and streams of northwest Michigan as the 75% of dead ash that are still standing fall. Such debris provides nutrients and habitat for an array of plants and animals, thereby influencing the abundance, activity and species compositions of several ground dwelling insects and seedling establishment. In streams, coarse woody debris provides complex habitat and refuges. It also retains organic matter. Recreationists do find that debris impedes boating.
Loss of ash specifically
As described by Engelken, Benbow and McCullough (2020), and in my earlier blog, ash leaf litter – particularly black ash leaf litter – is highly nutritious. Ash leaf litter has efficient turnover rates and contributes important soil nutrients such as nitrogen, organic carbon and exchangeable cations. Invertebrate communities in headwater streams feed largely on coarse organic material such as leaf litter (Engelken and McCullough 2020). Consequently, loss of the annual influx of ash leaf litter will likely have adverse effects on nutrient availability in riparian forests and adjacent streams.
SOURCES
Engelken, P.J., M.E. Benbow, D.G. McCullough. 2020. Legacy effects of emerald ash borer on riparian forest vegetation and structure. Forest Ecology and Management 457 (2020) 117684
Engelken, P.J. and D.G. McCullough. 2020. Riparian Forest Conditions Along Three Northern Michigan Rivers Following Emerald Ash Borer Invasion. Canadian Journal of Forest Research.
Siegert, N.W., P.J. Engelken, D.G. McCullough. 2021 Changes in demography and carrying capacity of green ash and black ash ten years after emerald ash borer invasion of two ash-dominant forests. Forest Ecology and Management Vol 494, August 2021
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
brown tree snake Boiga irregularis; via Wikimedia; one of the species on which the most money is spent on preventive efforts
In recent years a group of scientists have attempted to determine how much invasive species are costing worldwide. See Daigne et al. 2020 here.
Some of these scientists have now gone further in evaluating these data. Cuthbert et al. (2022) [full citation at end of blog] see management of steadily increasing numbers of invasive, alien species as a major societal challenge for the 21st Century. They undertook their study of invasive species-related costs and expenditures because rising numbers and impacts of bioinvasions are placing growing pressure on the management of ecological and economic systems and they expect this burden to continue to rise (citing Seebens et al., 2021; full citation at end of blog).
They relied on a database of economic costs (InvaCost; see “methods” section of Cuthbert et al.) It is the best there is but Cuthbert et al. note several gaps:
Only 83 countries reported management costs; of those, only 24 reported costs specifically associated with pre-invasion (prevention) efforts.
Data comparing regional costs do not incorporate consideration of varying purchasing power of the reporting countries’ currencies.
Data available are patchy so global management costs are probably substantially underestimated. For example, forest insects and pathogens account for less than 1% of the records in the InvaCost database, but constitute 25% of total annual costs ($43.4 billion) (Williams et al., in prep.) .
Still, their findings fit widespread expectations.
These data point to a total cost associated with invasive species – including both realized damage and management costs – of about $1.5 trillion since 1960. North America and Oceania spent by far the greatest amount of all global money countering bioinvasions. North America spent 54% of the total expenditure of $95.3 billion; Oceania spent 30%. The remaining regions each spent less than $5 billion.
Cuthbert et al. set out to compare management expenditures to losses/damage; to compare management expenditures pre-invasion (prevention) to post-invasion (control); and to determine potential savings if management had been more timely.
Economic Data Show Global Efforts Could Be – But Aren’t — Cost-Effective
The authors conclude that countries are making insufficient investments in invasive species management — particularly preventive management. This failure is demonstrated by the fact thatreported management expenditures ($95.3 billion) are only 8% of total damage costs from invasions ($1.13 trillion). While both cost or losses and management expenditures have risen over time, even in recent decades, losses were more than ten times larger than reported management expenditures. This discrepancy was true across all regions except the Antarctic-Subantarctic. The discrepancy was especially noteworthy in Asia, where damages were 77-times higher than management expenditures.
Furthermore, only a tiny fraction of overall management spending goes to prevention. Of the $95.3 billion in total spending on management, only $2.8 billion – less than 3% – has been spent on pre-invasion management. Again, this pattern is true for all geographic regions except the Antarctic-Subantarctic. The divergence is greatest in Africa, where post-introduction control is funded at more than 1400 times preventive efforts. It is also significant for Asia and South America.
Even in North America – where preventative actions were most generously funded – post-introduction management is funded at 16 times that of prevention.
Cuthbert et al. worry particularly about the low level of funding for prevention in the Global South. They note that these conservation managers operate under severe budgetary constraints. At least some of the bioinvasion-caused losses suffered by resources under their stewardship could have been avoided if the invaders’ introduction and establishment had been successfully prevented.
While in the body of the article Cuthbert et al. seem uncertain about why funding for preventive actions is so low, in their conclusions they offer a convincing (to me) explanation. They note that people are intrinsically inclined to react when impact becomes apparent. It is therefore difficult to motivate proactive investment when impacts are seemingly absent in the short-term, incurred by other sectors, or in different regions, and when other demands on limited funds may seem more pressing. Plus efficient proactive management will prevent any impact, paradoxically undermining evidence of the value of this action!
Aedes aegypti mosquito; one of the species on which the most money is spent for post-introduction control; photo by James Gathany; via Flickr
Delay Costs Money
The reports contained in the InvaCost database indicate that management is delayed an average of 11 years after damage was first been reported. Cuthbert et al. estimate that these delays have caused an additional cost of about $1.2 trillion worldwide. Each $1 of management was estimated to reduce damage by $53.5 in this study. This finding, they argue, supports the value of timely invasive species management.
They point out that the Supplementary Materials contain many examples of bioinvasions that entail large and sustained late-stage expenditures that would have been avoided had management interventions begun earlier.
Although Cuthbert et al. are not as clear as I would wish, they seem to recognize also that stakeholders’ varying perceptions of whether an introduced species is causing a detrimental “impact” might also complicate reporting – not just whether any management action is taken
Cuthbert et al. are encouraged by two recent trends: growing investments in preventative actions and research, and shrinking delays in initiating management. However, these hopeful trends are unequal among the geographic regions.
Which Taxonomic Groups Get the Most Money?
About 42% of management costs ($39.9 billion) were spent on diverse or unspecified taxonomic groups. Of the costs that were taxonomically defined, 58% ($32.1 billion) was spent on invertebrates [see above re: forest pests]; 27% ($14.8 billion) on plants; 12% ($6.7 billion) on vertebrates; and 3% ($1.8 billion) on “other” taxa, i.e. fungi, chromists, and pathogens. For all of these defined taxonomic groups, post-invasion management dominated over pre-invasion management.
When considering the invaded habitats, 69% of overall management spending was on terrestrial species ($66.1 billion); 7% on semi-aquatic species ($6.7 billion); 2% on aquatic species ($2.0 billion); the remainder was “diverse/unspecified”. For pre-invasion management (prevention programs), terrestrial species were still highest ($840.4 million). However, a relatively large share of investments was allocated to aquatic invaders ($624.2 million).
Considering costs attributed to individual species, the top 10 targetted for preventive efforts were four insects, three mammals, two reptiles, and one alga. Top expenditures for post-invasion investments went to eight insects [including Asian longhorned beetle], one mammal, and one bird.
Asian longhorned beetle
Just two of the costliest species were in both categories: insects red imported fire ant(Solenopsis invicta) and Mediterranean fruitfly (Ceratitis capitate). None of the species with the highest pre-invasion investment was among the top 10 costliest invaders in terms of damages. However, note the lack of data on fungi, chromists, and pathogens. (I wrote about this problem in an earlier blog.)
Discussion and Recommendations
Cuthbert et al. conclude that damage costs and post-invasion spending are probably growing substantially faster than pre-invasion investment. Therefore, they call for a stronger commitment to enhancing biosecurity and for more reliance on regional efforts rather than ones by individual countries. Their examples of opportunities come from Europe.
Drawing parallels to climate action, the authors also call for greater emphasis on during decision-making to act collectively and proactively to solve a growing global and inter-generational problem.
Cuthbert et al. focus many of their recommendations on improving reporting. One point I found particularly interesting: given the uneven and rapidly changing nature of invasive species data, they think it likely that future invasions could involve a new suite of geographic origins, pathways or vectors, taxonomic groups, and habitats. These could require different management approaches than those in use today.
As regards data and reporting, Cuthbert et al. recommend:
1) reducing bias in cost data by increasing funding for reporting of underreported taxa and regions;
2) addressing ambiguities in data by adopting a harmonized framework for reporting expenditures. For example, agriculture and public health officials refer to “pest species” without differentiating introduced from native species. (An earlier blog also discussed the challenge arising from these fields’ different purposes and cultures.)
3) urging colleagues to try harder to collect and integrate cost information, especially across sectors;
4) urging countries to report separately costs and expenditures associated with different categories, i.e., prevention separately from post-invasion management; damage separately from management efforts; and.
5) creating a formal repository for information about the efficacy of management expenditures.
While the InvaCost database is incomplete (a result of poor accounting by the countries, not lack of effort by the compilers!), analysis of these data points to some obvious ways to improve global efforts to contain bioinvasion. I hope countries will adjust their efforts based on these findings.
Seebens, H. S. Bacher, T.M. Blackburn, C. Capinha, W. Dawson, S. Dullinger, P. Genovesi, P.E. Hulme, M.van Kleunen, I. Kühn, J.M. Jeschke, B. Lenzner, A.M. Liebhold, Z. Pattison, J. Perg, P. Pyšek, M. Winter, F. Essl. 2021. Projecting the continental accumulation of alien species through to 2050. Glob Change Biol. 2021;27:970-982.
Williams, G.M., M.D. Ginzel, Z. Ma, D.C. Adams, F.T. Campbell, G.M. Lovett, M. Belén Pildain, K.F. Raffa, K.J.K. Gandhi, A. Santini, R.A. Sniezko, M.J. Wingfield, and P. Bonello 2022. The Global Forest Health Crisis: A Public Good Social Dilemma in Need of International Collective Action. submitted
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Hakalau Forest, Hawai“i; nearly 90% of Hawaiian flora is unique to the Islands
A recent article by Yang et al. 2021 (full citation at the end of this blog) seeks to determine the extent to which introduced plants reduce the uniqueness of regional floras. They analyzed data from 658 regions covering about 65.7% of the Earth’s ice-free land surface and about 62.3% of the planet’s known plant species.
They found strong homogenization of plant species’ taxonomic and phylogenetic diversity results from introductions of plant species to ecosystems beyond their native range. Homogenization caused by regional extinctions of native floral species occurs much less frequently.
There are two aspects of a region’s floral uniqueness. One is the number of species that it shares with other regions. This is taxonomic uniqueness. The other is the distinctiveness of the evolutionary history of the region. When several species are endemic to a region’s flora, and lack close relatives in other regions, that equals phylogenetic uniqueness.
The effect of a species introduction differs depending on which of these aspects one focuses on. Thus, naturalization of a species closely related to native species (e.g., a congeneric species) will have less impact on the phylogenetic floristic uniqueness of the region than naturalization by a distantly related species. Taxonomic uniqueness, however, will be affected to the same degree, irrespective of the phylogenetic distance between the naturalized and native species.
Yang et al. found strong homogenization of plant diversity. They found that species introductions increased the taxonomic similarity in 90.7% of all regional pairs and phylogenetic similarity in 77.2% of all region pairs. Most homogenization results from introductions of plant species to ecosystems beyond their native range. Homogenization caused by regional extinctions of native floral species occurs much less frequently.
This loss of regional biotic uniqueness or distinctiveness changes biotic interactions and species assemblages. These, in turn, have ecological and evolutionary consequences at larger scales and higher levels.
The degree of homogenization between regions’ floras depends on three factors:
1) The distance between the donor and recipient regions. Since nearby regions share more species, an introduction from a more distant origin is more likely to be a novel species and so contribute to homogenization of “donor” and “receiving” floras.
2) Climatic similarity, especially temperature. A plant species introduced from a climatically similar but geographically distant place is more likely to establish than a species from a different climatic zone. As a result, the recipient area’s flora is changed to more closely resemble the flora of the donor region with which it shares climatic conditions – regardless of the distance between them.
3) The level of exchange of goods and people between two regions. The higher the rate of exchange between two regions, the greater the chance that a species will be introduced and become established. Yang et al. used the existence of current or past administrative relationships (e.g., colonial relationship) between two regions as a proxy for intensity of trade and transport between donor and recipient regions. They found that floras of regions with current or past administrative links have taxonomically become more similar to each other than the floras of regions with no such links.
flora of the Cape Floral Kingdom – South Africa; photo from Michael Wingfield
Establishment of introduced species can increase floristic similarity of the donor and recipient regions (= floristic homogenization) when the species is native to one of the two regions and naturalizes in the other, or when it is not native to both regions and naturalizes in both. On the other hand, a species introduction can decrease the floristic similarity of the two regions (i.e., enhance floristic differentiation) when the species is not native to both regions but naturalized in only one.
Homogenization hotspots differed slightly depending on whether one focused on taxonomic or phylogenetic aspects.
The regions with the greatest average increase in taxonomic similarity with other regions due to naturalized alien species were New Zealand, portions of Australia, and many oceanic islands. The Australasian situation probably reflects its long biogeographic isolation from other parts of the globe and its highly unique native flora. As a result, nearly all non-native plants introduced to Australasia strongly increase levels of its floristic similarity to the rest of the world. Oceanic islands have species-poor floras with large proportions of unique endemics. They have also received high numbers of naturalized alien plants.
Hotspots of phylogenetic homogenization on continents are the same as those for taxonomic homogenization, but this is not true for islands. Yang et al. think this is because islands’ native floras were established by natural colonization from nearby continental floras so – despite subsequent speciation – they retain their phylogenetic relationship to the donor areas’ floras.
Yang et al. concede that they lacked high-quality data on native and naturalized alien species lists for a third of Earth’s ice-free terrestrial surface, especially Africa, Eastern Europe, and tropical Asia. They believe, however, that data from these regions are unlikely to change the overall finding. (Scientists are beginning to compile lists of forest pests in Africa). link to blog
Yang et al. note that introduction and naturalization of alien species are likely to increase in the future, thusaccelerating floristic homogenization. The ecological, evolutionary and socioeconomic consequences are largely unknown.They call for stronger biosecurity regulations of trade and transport and other measures to protect native vegetation.
SOURCE
Yang, Q., P. Weigelt, T.S. Fristoe, Z. Zhang, H. Kreft, A. Stein, H. Seebens, W. Dawson, F. Essl, C. König, B. Lenzner, J. Pergl, R. Pouteau, P. Pyšek, M. Winter, A.L. Ebel, N. Fuentes, E.L.H. Giehl, J. Kartesz, P. Krestov, T. Kukk, M. Nishino, A. Kupriyanov, J.L. Villaseñor, J.J. Wieringa, A. Zeddam, E. Zykova and M. van Kleunen. 2021. The global loss of floristic uniqueness. NATURE COMMUNICATIONS (2021) 12:7290. https://doi.org/10.1038/s41467-021-27603-y
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
EAB-killed ash tree lying on highway in Fairfax County, Virginia; photo by F.T. Campbell
A recent study (Hudgins, Koch, Ambrose & Leung 2022; full citation at end of blog) projects that, by 2050, 1.4 million street trees in urban areas and communities will be killed by introduced insect pests. This represents 2.1- 2.5% of all urban street trees. Nearly all of this mortality will occur in a quarter of the 30,000 communities evaluated. Additional urban trees – in parks, other plantings, on homeowners’ properties, and in urban woodlands – are also expected to die.
Loss of these trees will undercut all the ecosystem services provided by urban trees.
The principal cause of mortality will be the emerald ash borer (EAB). Already, an estimated 230,000 ash trees have been killed by EAB. The authors predict that 6,747 communities not yet affected by the EAB will suffer the highest losses between now and 2060. Most of these communities are in a 350,000 square mile area of the northeast and central states. However, the risk is far wider, reaching as far as Seattle.
This ash tree has been standing – dead – since 2016. When will it fall?
In the top ‘hotspot cities’ projected mortality is in the range of 5,000–25,000 street trees. These include Milwaukee; the Chicago area (Chicago / Aurora / Naperville / Arlington Heights); Cleveland; and Indianapolis. As in previous studies, the highest insect impacts are in the Northeast. Pests impacting this region – in addition to the emerald ash borer – include the spongy moth (formerly called gypsy moth) and hemlock woolly adelgid.
Because insect-killed trees must be treated or removed to minimize the risk to human life and property, the pest risk represents an economic as well as ecological threat. Removing and replacing just the street trees is projected to cost cities $30 million per year. Considering the cities I mentioned above, Milwaukee faces costs estimated at $13 million; Warwick, RI $2.5 million; Baltimore $1.7; Richmond and Virginia Beach $7.3 million and $700,000 respectively; and three New Jersey cities (Jersey City, Elizabeth City, and Patterson) $1.6 million combined.
USDA APHIS ended the federal quarantine for EAB in 2021. Therefore these cities and states are on their own to protect themselves from not only this and other damaging insects but also their extraordinarily high economic costs.
The study evaluated the risk to 48 genera of trees in about 30,000 communities. The most widely planted genera are maples (Acer spp.) and oaks (Quercus spp.). Consequently, they will die in largest numbers. An estimated 26.5 million maples and 5.9 million oaks are at risk, primarily in the East. As noted above, EAB is expected to kill 99% of ash trees in 6,747 communities across the country. In the Southwest, there are 3.4 million pines (Pinus spp.); the threat to them is not woodborers, but scale insects (San Jose scale [Quadraspidiotus perniciosus] and calico scale [Eulecanium cerasorum]).
As we know, urban forests are easily invaded because they are close to ports of entry and are often composed primarily of highly susceptible species. Hudgins, Koch, Ambrose and Leung analyzed the potential risk associated with introduction of a new woodboring insect from Asia – which they point out is the source of most imported goods. They determined that if such an introduced pest were to attack maples or oaks, it could kill 6.1 million trees and cost American cities $4.9 billion over 30 years. The risk would be highest if this pest were introduced via a port in the South.
In an earlier blog I reported that the U.S. is currently importing about 20 million shipping containers filled with goods from Asia per year. I have often blogged about the pest risk associated with wood packaging accompanying these imports. The number of containers from Asia entering Southeastern ports rose by more than 10% from December to January.
Hudgins, Koch, Ambrose & Leung combined four sources of information to produce these estimates:
a model of spread for 57 species of introduced insect pests already determined to cause significant damage to trees;
the distribution of genera of urban street trees across 30,000 US communities;
a model of host mortality in response to each insect-host combination; and
the cost of removing and replacing dead trees, linked to tree size (dbh).
They excluded several categories of pests. One of the most damaging, Asian longhorned beetle, was excluded because scientists have already developed control methods to limit its spread. Also excluded were species present in the U.S. for less than five years; species with no known economic impacts; and species for which no hosts in natural North American forests have been identified. Also excluded – although the authors do not mention this – are species that did not qualify for inclusion in the Aukema et al. study (see reference at end of this blog) because they have been introduced from nearby portions of North America, e.g., goldspotted oak borer. Finally, the study does not include pathogens. Some pathogens have caused huge losses of urban trees in the past, e.g., Dutch elm disease; some are causing losses now, e.g., sudden oak death. The authors do mention the Fusarium disease vectored by polyphagous (and Kuroshio) shot hole borers in southern California.
elm-lined street; photo from USFS
Consequently, the study’s estimate of 1.4 million street trees dead and costs of $30 million per year are underestimates.
The study has generated considerable media interest, including in the Washington Post.
SOURCES
Aukema, J.E., D.G. McCullough, B. Von Holle, A.M. Liebhold, K. Britton, & S.J. Frankel. 2010. Historical Accumulation of Nonindigenous Forest Pests in the Continental United States. Bioscience. December 2010 / Vol. 60 No. 11
Hudgins, E.J., F.H. Koch, M.J. Ambrose, B. Leung. 2022. Hotspots of pest-induced US urban tree death, 2020–2050. Journal of Applied Ecology 2022;00:1-11 DOI: 10.1111/1365-2664.14141
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
I report here on recent developments on breeding resistant trees. These include both promising results from decades-long efforts and also a promising start to addressing a new challenge.
These programs have benefited from major commitments by the USDA Forest Service. I hope they encourage similar commitments for other priority species – such as those named by the CAPTURE program.
Port-Orford cedar – ready to be planted in the forest!
Scientists who have been working for decades to breed seedlings of Port-Orford cedar (POC) trees resistant to the root rot caused by Phytophthora lateralis https://www.dontmovefirewood.org/pest_pathogen/port-orford-cedar-root-disease-html/now say that they have seedlings ready for planting in the forest. They made this case in a webinar in late February. It can be viewed here. The full webinar runs somewhat over two hours.
The scientist who led early studies of POC and the root disease, Don Zobel, Professor Emeritus, Oregon State University, described the ecological requirements that should guide planting programs. POC produces high-calcium litter. It grows from the sea coast to 1950 meters elevation, on sand dunes, fens, soils with hardpans; mafic & ultramafic rocks (serptentines) and fertile soils on some sedimentary rocks. POC is less shade tolerant than western hemlock but more fire tolerant. It can form a secondary canopy under Douglas-fir and supercede other conifers when fire occurs repeatedly. The tree needs surface water, e.g., seepages and stream sides; but the water must be flowing, not stagnant. Seedlings are especially vulnerable to drying during winter.
[I posted a separate blog about other trees native to this region, including serpentine soils, here.]
One purpose of the webinar was to encourage owners and managers of lands within POC’s historic range (see the map under Dr. Zobel’s presentation) to begin planting the species in appropriate sites. With this in mind, Dr. Zobel emphasized criteria for selecting sites:
Climates in coastal areas of the range are less likely to change under climate change
Quartenary marine terraces are the best geologic type; Lookingglass and Roseburg geologic types are also acceptable
Availability of water during summer, e.g., streamside and seepage areas. Try planting beneath alder. However, avoid interior valley stream corridors if the soils are not ultramafic. And avoid stagnant water.
a POC tree in a bog next to the endemic pitcher plant of southern Oregon, Darlingtoniacalifornica; photo by Richard Sniezko
Dr. Zobel also says one should plant pathogen-resistant genotypes and pay attention to local genetic varieties (which have largely been determined).
Dr. Richard Sniezko of the USFS Dorena Genetic Resource Center described the Center’s 30-year effort to find and exploit resistance to the pathogen. Funding has come from the USFS Forest Health Protection program, other parts of the USFS, and the Bureau of Land Management (BLM). The goal all along has been to produce seedlings for restoration to the forest – meaning not just resistant to the pathogen but also adapted to various local conditions. The program can now provide resistant seedlings in large quantities for planting by landowners and public land managers.
Dr. Sniezko emphasizes that success depends on engagement of four sets of people: research by university scientists; application of that research and development of propagule growing methods by the Dorena Center; support from USFS leaders to continue the program; involvement of land managers who choose to plant the resistant seedlings.
USFS and BLM staff described efforts to determine where POC grows on land under their management, the status of disease in those areas, and efforts to slow the spread of the disease, especially along roadsides and as result of timber or engineering projects. Some of this sanitation work has been funded by USFS Forest Health Protection program — not the National Forest System.
Richard Sniezko stated that the seedlings’ quantitative disease resistance means that some seedlings will die. He expects 40-50% survival of seedlings from many of the breeding zones. This is well above the level of resistance in un-improved populations.
Both BLM and the Rogue-River-Siskiyou National Forest have planted tens of thousands of resistant seedlings in recent years and plan to continue. Funding provided by COVID-19 legislation might allow increased effort. [See Dr. Sniezko’s presentation on the webinar for photos from some plantings.]
POC seedlings at Dorena; photo by Richard Sniezko
Norma Kline of the Oregon State University extension program has distributed more than 10,000 seedlings to small/non-industrial landowners. Many of the recipients shared seedlings with neighbors or are coordinating their planting over a large area. They were motivated primarily by conservation concerns. Her monitoring showed that the POC seedlings survived but did not thrive under dense tanoak canopy. They did well in competition with grass in areas near the coast where there was more moisture. They also did well under Douglasfir as long as there was dappled sunlight.
The non-governmental organization American Forests is likely to participate actively in the planting effort.
In an email to me, Dr. Sniezko asks that people who have planted POC outside its native range inform him where the tree(s) is/are thriving. This information would enhance scientists’ understanding of the species’ environmental tolerances.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
‘Ohi‘a (Metrosideros polymorpha) is the most abundant native forest tree in Hawai`i and of enormous ecological, cultural, and economic importance. Five species of endemic Metrosideros are recognized on the Hawaiian islands. Only one — M. polymorpha — is found throughout the state. Eight varieties are recognized. These varieties inhabit different environments and have adapted to selective pressures characteristic of these locations. There are at least five other species in the Metrosideros genus, each endemic to one or a few nearby islands. Blaine et al. (2022) [full citation at end of this blog] provide a helpful summary of the tree’s ecological importance and its apparently on-going speciation.
‘Ohi‘a provides habitat for endemic birds, insects, and plants, many of which are endangered. Thus, conservation of this species — and all Hawaiian Metrosideros – is vital for the conservation of countless other taxa. In addition, high elevation ʻōhiʻa forests protect vitally important watersheds across the state. For Native Hawaiians, ʻōhiʻa is a physical manifestation of multiple Hawaiian deities so is the subject of many proverbs, chants, stories, and a foundation of scared hula. Finally, the tree is beautiful!
Native Hawaiian forests face multiple threats — invasive animals and plants, wildfire, and land-use changes. Due to such threats, natural ʻōhiʻa regeneration is largely absent in most lower-elevation forests. In this case, competition with invasive species and the presence of diseases such as ʻōhiʻa rust (Austropuccinia psidii) are probably the specific causes. Multiple government and non-governmental entities have made substantial effort to mitigate these threats.
ROD-infected ʻōhiʻa; photo by J.B. Friday
The disease Rapid ‘Ohi‘a Death (ROD) is an unprecedented threat to this species and the forests it constitutes. The disease is caused by two newly described fungal pathogens: Ceratocystis lukuohia and C. huliohia. The disease caused by C. lukuohia is more severe. To date it has been detected on the two islands farthest apart in the chain — Hawai`i (the Big Island) and Kaua‘i. C. huliohia causes a canker disease that kills trees more slowly. It is more widespread, found on Maui and O‘ahu in addition to Hawai`i and Kaua‘i. Blaine et al. (2022) and the profile here describe the two diseases’ epidemiologies, progression, impacts, and challenges.
Because of the clear threat to Hawaiian ecosystems, ecosystem services, and cultural assets, considerable effort has put into delimitation and research on possible mitigation actions since ROD was discovered in 2010. The first strategic plan covered the period 2017–2019. It focused on expanded efforts to map outbreaks, research on the epidemiology of the pathogens, and most-promising management practices. The second strategic plan covers 2020–2024. It provides for continued surveillance and improvement of these technologies; expanding outreach and public engagement; research on possible vectors of the pathogens; collection and preservation of seeds for research and future restoration; and comprehensive evaluation and development of disease resistance in ʻōhiʻa.
Soon after the causal agents were clarified, the USDA Agriculture Research Service (ARS) began screening for disease resistance. By 2016, ARS had demonstrated that five individuals from two varieties of M. polymorpha had survived inoculation by the more virulent pathogen, C. lukuohia. Their survival raised hopes that natural resistance might be present in wild populations of at least some varieties. However, more comprehensive screening of trees from throughout the species’ range is needed to provide an accurate baseline on the frequency, level, and distribution of genetic resistance to both pathogens. The goal is to produce material resistant to both pathogens that can be used to preserve the ecology, culture, and biotic communities that are dependent on this tree species.
To carry the expanded effort forward, in 2018 a collaborative partnership of state, federal, and non-profit groups was formed. Participants in the ‘Ohi‘a Disease Resistance Program (‘ODRP) include: the Akaka Foundation for Tropical Forests; USDA’s Forest Service and Agriculture Research Service; the state’s Division of Forestry and Wildlife and Agriculture Research Center; programs of the University of Hawai‘i at Manoa and at Hilo; Purdue and Arizona State universities; the Tropical Hardwood Tree Improvement and Regeneration Center; and Kalehua Seed Conservation Consulting.
Blaine et al. (2022) have now outlined a framework to guide the overall effort to identify and develop ROD resistance in M. polymorpha and, possibly, all Hawaiian Metrosideros species. The framework calls for the following activities:
(1) evaluating and operationalizing methods for inoculation-based screening and greenhouse-based production of test plants; and
(2) short-term greenhouse screenings of seedlings and rooted cuttings sampled from native Metrosideros throughout Hawai’i.
Once these tasks have been achieved, the effort is expected to expand to address:
(3) establishing field trials to validate the short-term greenhouse assays and monitor durability and stability of resistance;
drivers of susceptibility and resistance to characterize the durability and stability of genetic resistance to ROD;
(5) developing remote sensing and molecular methods to rapidly detect ROD-resistant individuals;
(6) if necessary, conducting breeding to increase the efficacy of resistance and improve durability of ROD resistance; and
(7) supporting already established and ongoing Metrosideros conservation, including state-wide seed collection and banking, with information on not only genotypes resistant to ROD but also production of ROD-resistant seed.
Blaine et al. (2022) outline how to proceed on each step, and describe the challenges that must be overcome. Challenges range from building growing and screening capacity to handle the thousands of plants required, to developing the remote sensing tools to identify diseased trees in the forest, to identifying sites for seed orchards. Actions by ‘ODRP will focus on Stage II screening in the field to examine the durability of resistance under the wide variety of ecological conditions in which ʻōhiʻa grows and in the presence of a potentially evolving pathogen. Resistance studies must expand beyond M. polymorpha varieties from only one island (the Big Island) to include the other Hawaiian Metrosideros taxa.
Once ROD-resistant M. polymorpha trees are discovered and groundwork has been laid to satisfy initial needs for resistant tree seedlings for forest restoration, scientists can begin research into the genetic basis of ROD resistance. This knowledge will assist breeding efforts which might be necessary if resistance to one of the pathogens does not confer resistance to the other, since the goal is to provide seedlings that are resistant to both.
Blaine et al. (2022) note that the state and others continue efforts to address other aspects of ROD management. These include
1) controlling the spread of the pathogen through local quarantines on movement of infected material and increased public education on bio-sanitation for forest users;
2) testing repellants to reduce beetle attack on infected trees and subsequent frass production.
3) reducing wounding of trees by fencing more pristine forests and removing feral ungulates
SOURCE
Blaine C. Luiz, Christian P. Giardina, Lisa M. Keith, Douglass F. Jacobs, Richard A. Sniezko, Marc A. Hughes, James B. Friday, Philip Cannon, Robert Hauff, Kainana Francisco, Marian M. Chau, Nicklos Dudley, Aileen Yeh, Gregory Asner, Roberta E. Martin, Ryan Perroy, Brian J. Tucker, Ale.alani Evangelista, Veronica Fernandez, Chloe Martins-Keli.iho.omalu, Kirie Santos, Rebekah Ohara. 2022. A framework for establishing a rapid ‘Ohia death resistance program. New Forests. https://doi.org/10.1007/s11056-021-09896-5
See also the video at https://www.bigislandvideonews.com/2019/06/16/video-to-save-ohia-a-genetic-resistance-program-will-be-built/
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
vegetation killed by Phytophthora cinnamomi in West Australia
Some invasive species practitioners have been trying to develop a standardized framework for describing bioinvasions. Their goal is to overcome disparities in approaches developed by scientists working with various taxonomic groups in hopes of improving understanding of, and communication about, bioinvasions. Prominent among these efforts is the “Unified Framework for Bioinvasion” published by Blackburn et al. in 2011 (full citation at end of blog).
Now several forest pathologists (Paap et al; full citation at end of blog) say that this framework does not adequately integrate forest pathogens. This omission is particularly unfortunate given the prominence of forest pathogens as damaging invaders – e.g., chestnut blight in Europe and North America; white pine blister rust in North America; sudden oak death in North America and Great Britain; myrtle rust and Phytophthora cinnamomi in Australia. (See profiles of all these pathogens here; I note additional examples in North America, such as laurel wilt disease.)
Paap et al think that this omission impedes understanding of both forest pests and invasive species in general. Also, they say that integrating microorganisms into the broader Blackburn framework would help forest pathologists better understand how and why invasions occur, where they occur, and how they can be stopped or mitigated.
Furthermore, they note the importance of integrating the diverging terminologies used by invasive species practitioners and plant pathologists and their separate regulatory bodies – the Convention on the Conservation of Biological Diversity (CBD) and the International Plant Protection Convention (IPPC). I concur, since nations’ programs regulating plant diseases and their vectors operate under the IPPC rubric.
Figure 2 and Table 1 lay out Paap et al.’s proposed modification of Blackburn’s framework, and detail strategies linked to management goals appropriate for the stages of plant disease development.
Tanoak mortality in southern Oregon caused by P. ramorum – a pathogen completely unknown until it was introduced to North America and Europe; photo by Oregon Department of Forestry
However, such integration will be impeded by many difficulties (I have re-ordered these points):
1) The first – which underlies all others — is the paucity of data on microbial taxa, which undermines the pest risk analyses and other systems developed for assessing and managing other types of invasive species. That is,
Many of the vast number of microbial taxa have not yet been described.
Even species that have been describe often cannot be ascribed to a specific geographic origin. This information gap undercuts efforts to determine whether a disease outbreak is caused by an “introduced” organism.
2) Microbial species are usually detected only when disease impacts become obvious. However, an outbreak might not signal a new or spreading “introduction”. While invasive species must—by definition—cross a geographic boundary (through the assistance of human actions), pathogens can cause disease outbreaks through breaching a wider range of boundaries, including ecological and evolutionary ones. Thus, the disease outbreak doesn’t always fit the definition of “invasive species”.
3) Substantial differences exist in training and goals between fields. Forest pathologists are usually trained in plant pathology (often focused on crops) rather than in forestry or ecology. Their goal is to manage the pathogen. Invasion scientists tend to focus on natural ecosystems, study animal and plant invasions, and seek understanding of the invasion process.
4) A related issue is that the two fields operate under separate regulatory bodies that have different emphases and aims. Paap et al. note that while the IPPC ostensibly includes impacts on natural environments, its members’ priority is plants of economic importance. The World Trade Organization’s Agreement on the Application of Sanitary and Phytosanitary Measures (WTO SPS) seeks primarily to minimize disruption of trade resulting from plant health regulation. On the other hand, the CBD explicitly considers invasive species’ impact to the natural environment (Aichi Biodiversity Target 9). [To read my critique of the WTO SPS and IPPC, read the Fading Forests reports (link at end of this blog), especially FF II.]
Rome – home to the IPPC
They note that in 2004, the IPPC and CBD secretariats established a Memorandum of Cooperation to promote synergy and to avoid duplication. Paap et al. appear disappointed that despite development of joint work plans, phytosanitary programs are still focused largely on crop pathogens.
Disease development – a complex set of circumstances that makes risk assessment less reliable
Since I am not a pathologist (or even a biologist), I learned a lot about the complexities of plant pathology from Paap et al.
While I am certainly familiar with the “disease triangle” concept, I had not thought about certain implications. For example, pathogens can cause severe disease outbreaks by evading any one of three types of barriers: geographic, environmental, or evolutionary. Transport of the micro-organism to a new ecosystem (leaping the geographic barrier and meeting the definition of an “introduction” in invasive species terminology) certainly can facilitate disease outbreaks. However, evolutionary and environmental barriers might also be overcome in other ways.
The result is that a plant disease can develop under multiple scenarios following the introduction of an alien pathogen. These scenarios are:
disease on a coevolved host growing as an alien species in the new environment, for example plantations of trees grown for timber (pathogen reunion);
disease on a naïve host that is itself alien to the geographic region in question (host jump);
disease on an alien host (naïve or coevolved) which supports disease on a host native to the new geographic area that could not be sustained in the absence of the alien host;
disease on alien and native hosts; and
disease on a host native to the new geographic area but not on an alien host.
Countries’ efforts to conduct pest risk analyses are unlikely to be straightforward – or even possible – with so many disease scenarios
Paap et al. proceed to compare introductory pathways under the CBD categorization and plant pathology. In doing so they point out several aspects of introduction, establishment, and spread that are specific to pathogens. For example, trees’ long life spans and inability to adapt as rapidly as the micro-organism increase their vulnerability to devastating disease outbreaks following the arrival of a novel pathogen.
Participants in the Montesclaros meeting that drafted an early critique of international phytosanitary procedures
Paap et al. reinforce points made by other critics of current phytosanitary programs. (See my earlier blogs under the category “plants as pest vectors”.) In particular, they point out the weakness of visual inspection and note that new molecular assays can detect only known microorganisms. An additional complication is that DNA can persist in soil and plant tissue after death of the organism, leading to false positives. RNA is cannot yet be used as a viability marker.
Paap et al. provide three case studies to illustrate in greater depth several major challenges encountered when managing invasive forest pathogens. Most of these weaknesses are well known to forest pathologists.
1. The inconspicuous nature of microorganisms
As noted by Paap et al. and other authors, the difficulty detecting microbes is exacerbated by the huge volumes of goods, especially live plants, in international trade; the small proportion of those plants that can be inspected; the weakness of visual examination; application of fungicides and fertilizers before export that suppress symptoms. The chosen example is the oomycete genus Phytophthora, specifically P. ramorum.
2. Cryptic status of many species
Current biosecurity programs rely on naming the organism and its place of origin. This is actually impossible for many microorganisms. The tardy response to ash dieback (Hymenoscyphus fraxineus) in Europe illustrates the delay in determining the causal agent and its geographic origin. During this nearly two-decade period the possibility of preventing spread was lost.
3. Rapid evolution
Rapid evolution of the introduced pathogen can overcome resistance in a host. The example described is Cronartium ribicola (causal agent of white pine blister rust) on Western white pine (Pinus monticola) and sugar pine (P. lambertiana). They also mention the threat from hybridization between previously isolated populations, specifically Phytophthora x alni causing a devastating decline of black alder in Europe.
Sugar pine in Sequoia National Park; photo by S. Rae via Flickr
Paap et al. call for increased research to increase our knowledge of microbial diversity, especially in taxonomically rich and poorly studied ecosystems. They praise sentinel plantings as a powerful tool for early warning of pathogen threats.
SOURCES
Blackburn, T.M., P. Pysek, S. Bacher, J.T. Carlton, R.P. Duncan, V. Jarosik, et al. A proposed unified framework for biological invasions. Trends Ecol Evol. 2011; 26(7):333-9.
Paap, T., M.J. Wingfield, T.I. Burgess, J.R.U. Wilson, D.M. Richardson, A. Santini. 2022. Invasion Frameworks: a Forest Pathogen Perspective. FOREST PATHOLOGY https://doi.org/10.1007/s40725-021-00157-4
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Eucalyptus plantation in Kwa-Zulu-Natal, South Africa; Kwa-Zulu-Natal Dept. of Transportation
Graziosi et al. (full citation at the end of the blog) point out that trees are crucial for Africa’s future. Eight hundred of the 4,500–6,000 indigenous tree species provide significant food. As elsewhere, trees provide wood and other extractive resources essential for economic growth. They also support biodiversity and mitigate current and impending climatic variations. Africa– especially the Sub-Saharan countries – is already considered highly vulnerable to climate change.
According to Graziosi et al., the cumulative economic impact of all invasive species in Africa is expected to exceed $1.2 billion per year. The total invasion cost as a proportion of GDP for many African countries is among the highest in the world. This raises the stakes for developing locally appropriate management strategies across the continent.
Responding effectively to this threat is hampered by gaps in data as well as some countries’ limited capacity for biosecurity. Graziosi et al. say that improved knowledge of taxonomy, distribution, and damage caused by these organisms is essential. Such knoledge will be crucial to develop continent-wide strategies to manage this emergency and to enhance capacity for country-level interventions.
Native and alien pests. Indigenous and plantation trees
Africa’s trees and their services are threatened by both native pests and accelerating introductions of pests and diseases from elsewhere. Long-established and new invaders increasingly affect planted forests of exotic eucalypts, pines, and Australian acacias, as well as important indigenous trees. Graziosi et al. note that the U.N. Food and Agriculture Organization (FAO) in an annex to a report issued in 2009 recorded about100 species of forest pests affecting trees in planted and natural forests across Africa. Half are native insects and pathogens, a third are alien; about 15% are of unknown origin. Considering all pests, broadleaf trees (predominantly native) are most affected.
The result is damage from the local – e.g., to rural livelihoods – to the continental – e.g., to economic development and biological diversity across Africa. Moreover, pests exacerbate widespread loss of forest cover. Overall, African forests are shrinking at the rate of almost 0.5% annually. This deforestation is affecting particularly natural forests; planted forests are actually growing 1.3% annually.
Exotic plantation trees face severe threats. More than 47 native and 19 non-indigenous defoliators, sap-feeders, wood- and shoot-borers attack plantations of Acacia spp., Eucalyptus spp., Pinus spp., and teak (Tectona grandis). About 90% of pathogens of plantation forestry are either non-indigenous or of uncertain origin. Eucalyptus alone are severely damaged by 15 species of pathogens. These organisms are listed in Tables 1 and 2.
Numerous native insect species, known as pests of indigenous trees, have reportedly widened their host range and now damage exotic trees too. Some introduced insects appear to pose significant threats to native tree species. One example is the Cypress aphid Cinara cupressi, which is attacking both exotic cypress plantations and the native African cedar Juniperus procera. Some fungi in the family Botryosphaeriaceae are latent pathogens infecting a wide range of hosts including indigenous Acacia. Dieback of large baobab trees was recently reported from southern Africa. While various microorganisms are associated with these symptoms, the specific cause is still uncertain.
A baobab tree in Limpopo region of South Africa; Wikimedia
The risk currently appears to be particularly high in South Africa. The country’s flora is highly diverse and has a high level of endemism. In fact, South Africa is home to the Earth’s smallest floral kingdom, the Cape Floral Kingdom. It is also the apparent hot spot for pest introductions from overseas (see below). Phytophthora cinnamomi is attacking native Proteaceae in South Africa. According to Graziosi et al., an “incredible diversity” of Phytophthora taxa is present, portending threats to additional plant species. Other pathogens are attacking native conifers in the Podocarpus genus, Ekebergia capensis (Meliaceae), and Syzygium trees.
Protea repens and fynbos vegetation near Table Mountain; photo by Mike Wingfield
There is a clear pattern to further spread: pests first introduced to South Africa often spread. Examples include several insects and pathogens on Eucalyptus and the wood-boring pest of pine Sirex noctilio. This pattern is explained by two main factors. South Africa has a high capacity to detect introduced species. Also, there is an active plantation forestry sector that imports propagules. This offers opportunities for contaminating organisms to be introduced simultaneously.
Furthermore, as Graziosi et al. note, determining the geographic origin of significant proportion of pathogens is extremely difficult – an issue I will discuss in a separate blog based on a publication by primarily South African scientists. Some non-indigenous pathogens have been on the African continent for a long time. The Armillaria root rot pathogen apparently was introduced to South Africa with potted plants from Europe in the 1600s! They note also that many non-indigenous pathogens are probably already established on the continent but not yet detected due to the organisms’ cryptic nature and lagging detection abilities.
The future of African forests
African countries expect economic growth with associated increased trade with countries off-continent. The probable result will be to accelerate the rate of species introductions and spread. However, as climate change worsens, managers will find it increasingly difficult both to predict introduced species’ impact and to implement management programs.
This led Graziosi et al. to call for urgent improvements in plant biosecurity across the continent. They advocate improved coordination at regional and international levels. The list of needed actions is a familiar one: development and application of improved diagnostic tools, updated plant exchange regulations, and revised trade policies.
Graziosi et al. also call for development of effective control and management options. They suggest biocontrol, innovative silviculture practices, and selection of resistant trees. The good news is that African countries have already initiated programs to conserve tree germplasm and domesticate indigenous species, including establishment of field gene banks of high-priority indigenous trees. I have previously praised South African efforts, specifically reports here and here.
Mudada, Mapope, and Ngezimana (2022) describe the risk from introduced species to agriculture and human well-being in southern Africa beyond forestry. The region is already ravaged by food insecurities and hidden hunger. It would be devastated if the global average of crop loss due to plant diseases (10-16%) occurs there. They say these losses can be avoided with improved biosecurity mechanisms focused primarily on pest exclusion and plant quarantine regulations.
SOURCES
Graziosi, I. M. Tembo, J. Kuate, A. Muchugi. 2020 Pests and diseases of trees in Africa: A growing continental emergency. Plants People Planet DOI: 10.1002/ppp3.31
Mudada, N. Mapope, N., and Ngezimana, W. 2022 – The threat of transboundary plant pathogens to agricultural trade in Southern Africa: a perspective on Zimbabwe’s plant biosecurity – A review. Plant Pathology & Quarantine 12(1), 1–33, Doi 10.5943/ppq/12/1/1
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter the United States and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm