African Tulip Tree (Spathodea campanulata) on Puerto Rico; photo by Joe Schlabotnik via Flickr
While it is widely accepted that tropical island ecosystems are especially vulnerable to invasions, there has been little attention to terrestrial bioinvaders in the Caribbean; there has been more attention to marine bioinvaders such as lionfish. I am glad that is starting to change. Here I review a new study by Potter et al. (full citation at end of this blog), supplemented by information from other recent studies, especially Poland et al.
Potter et al. used USFS Forest Inventory and Analysis (FIA) survey data to examine regeneration rates by non-native tree species introduced to the continental United States, Hawai`i, and Puerto Rico. I rejoice that they have included these tropical islands, often left out of studies. They are part of the United States and are centers of plant endemism!
Potter et al. sought to learn which individual non-indigenous tree species are regenerating sufficiently to raise concern that they will cause significant ecological and economic damage in the future. That is, those they consider highly invasive. They defined such species as those for which at least 75% of stems of that species detected by FIA surveys are in their small tree categories – saplings or seedlings. They concluded that these species are successfully reproducing after reaching the canopy so they might be more likely to alter forest ecosystem functions and services. They labelled species exhibiting 60 – 75% of stems in the “small” categories as moderately invasive.
The authors recognize that many factors might affect tree species’ regeneration success, especially at the stand level. They assert that successful reproduction reflects a suite of factors such as propagule pressure, time since invasion, and ability of a species to adapt to different environments.
As I reported in an earlier blog, link 17% of the total flora of the islands of the Caribbean archipelago – including but not limited to Puerto Rico – are not native (Potter et al.). In Puerto Rico, two-thirds of forests comprise novel tree assemblages. The FIA records the presence of 57 non-native tree species on Puerto Rico. Potter et al. identified 17 non-native tree species as highly invasive, 16 as potentially highly invasive, and two as moderately invasive. That is, 33 of 57 nonnative tree species, or 58% of those species tallied by FIA surveyors, are actual or potential high-impact bioinvaders. While on the continent only seven non-native tree species occurred on at least 2% of FIA plots across the ecoregions in which they were inventoried, on Puerto Rico 21 species occurred on at least 2% of the FIA plots (38%). They could not assess the invasiveness of the eight species that occurred only as small stems on a couple of survey plots. These species might be in the early stages of widespread invasion, or they might never be able to reproduce & spread.
The high invasion density probably reflects Puerto Rico’s small size (5,325 mi² / 1,379,000 ha); 500 years of exposure to colonial settlement and global trade; and wide-scale abandonment of agricultural land since the middle of the 20th Century
Naming the invaders
The most widespread and common of the highly invasive non-native tree species are river tamarind (Leucaena leucocephala), on 12.6% of 294 forested plots; algarroba (Prosopis pallida) on 10.9%; and African tuliptree (Spathodea campanulata)on 6.1%. Potter et al. attribute the prevalence of some species largely to land-use history, i.e., reforestation of formerly agricultural lands. In addition, some of the moderately to highly invasive species currently provide timber and non-timber forest products, including S.campanulata, L. leucocephala, Syzgium jambos (rose apple) and Mangifera indica (mango).
Potter et al. contrast the threat posed by Spathodea campanulata with that posed by Syzgium jambo. The latteris shade tolerant and can form dense, monotypic stands under closed canopies. Because it can reproduce under its own canopy, it might be able to remain indefinitely in forests unless it is managed. In contrast S. campanulata commonly colonizes abandoned pastures. Since it is shade intolerant, it might decline in the future as other species overtop it. Meanwhile, they suggest, S. campanulata might provide habitat appropriate for the colonization of native tree species.
Second-growth forest in Caribbean National Forest “El Yunque”
Poland et al. say the threat from Syzgium jambos might be reduced by the accidentally introduced rust fungus Puccinia psidii (= Austropuccinia psidii), which has been killing rose apple in Puerto Rico. In Hawai`i, the same fungus has devastated rose apple in wetter areas.
Potter et al. note that stands dominated by L. leucocephala and Prosopis pallida in the island’s dry forests are sometimes arrested by chronic disturbance – presumably fire. However, they do not report whether other species – native or introduced – tend to replace these two after disturbance. The authors also say that areas with highly eroded soils might persist in a degraded state without trees. The prospect of longlasting bare soil or trashy scrub is certainly is alarming.
Potter et al. warn that the FIA’s sampling protocol is not designed to detect species that are early in the invasion process. However, they do advise targetting eradication or control efforts on the eight species that occurred only as small stems on a couple of survey plots. While their invasiveness cannot yet be determined, these species might be more easily managed because presumably few trees have yet reached reproductive age. They single out Schinus terebinthifolius (Brazilian pepper), since it is already recognized as moderately invasive in Hawai`i. I add that this species is seriously invasive in nearby peninsular Florida and here! APHIS recently approved release of a biocontrol insect in Florida targetting Brazilian pepper. It might easily reach nearby Puerto Rico or other islands in the Caribbean. I am not aware of native plant species in the Caribbean region that might be damaged by the biocontrol agent. However, two native Hawaiian shrubs might be harmed if/when this thrips reaches the Hawaiian Islands. Contact me for specifics, or read the accompanying blog about Potter et al. findings in Hawai`i.
Poland et al. looked at the full taxonomic range of possible bioinvaders in forest and grassland ecosystems. The Caribbean islands receive very brief coverage in the chapter on the Southeast (see Regional Summary Appendices). This chapter contains a statement that I consider unfortunate: “Introduction of species has enriched the flora and fauna of Puerto Rico and the Virgin Islands.” The chapter’s authors assert that many of the naturalized species are restoring forest conditions on formerly agricultural lands. They say that these islands’ experience demonstrates that introduced and native species can cohabitate and complement one another. I ask – but in what kind of forest? These forests, are novel communities that bear little relationship to pre-colonial biodiversity of the islands. Was not this chapter the right place to note that loss? Forests are more than CO2 sinks.
I also regret that the chapter does not mention that the Continental United States can be the source of potentially invasive species (see several examples below).
Mealybug-infested cactus at Cabo Rojo National Wildlife Refuge, Puerto Rico. Photo by Yorelyz Rodríguez-Reyes
The chapter does concede that some introduced species are causing ecological damage now. See Table A8.1. Some of these troublesome introduced species are insects:
the South American Harrisia cactus mealybug (Hypogeococcus pungens) is killing columnar cacti in the islands’ dry forests. The chapter discusses impacts on several cactus species and control efforts, especially the search for biocontrol agents.
the agave snout weevil (Scyphophorus acupunctatus), native to the U.S. Southwest and Mexico , is threatening the endemic and endangered century plant (Agave eggersiana) in St. Croix & Puerto Rico.
Tabebuia thrips (Holopothrips tabebuia) is of unknown origin. It is widespread around mainland Puerto Rico. Its impacts so far are primarily esthetic, but it does apparently feed on both native and introduced tree species in the Tabebuia and Crescentia genera.
The Caribbean discussion also devotes welcome attention to belowground invaders, i.e., earthworms. At least one species has been found in relatively undisturbed cloud forests, so it is apparently widespread. Little is known about its impact; more generally, introduced earthworms can increase soil carbon dioxide (CO2) emissions as through speeded-up litter decomposition and soil respiration.
A factsheet issued by the British forestry research arm DEFRA reports that the pine tortoise scale Toumeyella parvicornis has caused the death of 95% of the native Caicos pine (Pinus caribaea var. bahamensis) forests in the Turks and Caicos Islands (a UK Overseas Territory). The scale is native to North America. It has recently been introduced to Italy as well as to Puerto Rico, and the Turks and Caicos Islands.
SOURCES
Lugo, A.E., J.E. Smith, K.M. Potter, H. Marcano Vega, C.M. Kurtz. 2022. The Contribution of Non-native Tree Species to the Structure & Composition of Forests in the Conterminous United States in Comparison with Tropical Islands in the Pacific & Caribbean. USFS International Institute of Tropical Forestry General Technical Report IITF-54.
Poland, T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019), Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Especially the Appendix on the Southeast and Caribbean. Springer Verlag. Available gratis at https://link.springer.com/book/10.1007/978-3-030-45367-1
Potter K.M., Riitters, K.H. & Guo. Q. 2022. Non-nativetree regeneration indicates regional & national risks from current invasions. Frontiers in Forests & Global Change Front. For. Glob. Change 5:966407. doi: 10.3389/ffgc.2022.966407
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
At CISP, our hearts go out to all those affected by the terrible August fires on Maui. May the departed rest in peace. May the living find comfort and all that is needed for recovery.
Fire and Invasive Grasses
A fire in non-native grasses on Maui in 2009; photo by Forrest and Kim Starr
Major U.S. and international media continue to detail the fires’ devastation, especially in Lahaina. As time has passed, more news has highlighted the role that the widespread presence of introduced, fire-prone grasses played in the rapid growth and spread of Maui’s fires.
For example, The Washington Post devoted seven paragraphs in one story to the issue of grasses. The story quotes several experts: Alison Nugent, an associate atmospheric scientist at the University of Hawaii’s Water Resources Research Center; Jeff Masters, a meteorologist for Yale Climate Connections; and Clay Trauernicht, a fire researcher at the University of Hawaii.
These and others have been widely quoted in the many recent articles. I am glad that they – and the media – are making clear that climate change is not the sole factor causing damaging wildfires. It is clear that Maui’s recent weather patterns – including the high-velocity winds and drought – have been within the range of normal climate patterns. Fluctuations in the Pacific’s weather have also been normal, especially under the influence of the current El Niño.
The dangers caused by Hawai’i’s fire-prone grasses are also clear – and have been for years. Experts have identified policy weaknesses at the county and state level. Also, they have specified changes to land management that could better prevent or mitigate wildfires. There has been far too little action.
On the other hand, there are hopeful signs.
endangered ‘akikiki photo by Carter Atkinson, USGS
The Hawai’i Wildfire Management Organization, a nonprofit, is educating and engaging communities state-wide. Elizabeth Pickett, a Co-Executive Director, presented an overview of wildfire at the Hawai’i Invasive Species Awareness Month in February 2023. The Big Island Invasive Species Committee has successfully eradicated two species of pampas grass on Hawai’i Island – after 13 years’ work. A native species has been planted where pampas formerly grew.
Another Postarticle reported on efforts by staff and fire departments to protect the Maui Bird Conservation Center, which houses critically endangered Hawaiian birds found nowhere else on Earth, including some currently extinct in the wild. As I have blogged previously, the palila, kiwikiu, ‘akikiki, ‘alalā [Hawaiian crow; extinct in the wild] and other birds are dying from avian malaria, carried by nonnative mosquitoes. The Center on Maui and another on the Big Island are run by the San Diego Zoo Wildlife Alliance. Conservationists have completed field trials of a proposed mosquito suppression process for Maui and are seeking public comments for a similar program on Kaua’i. These programs represent groundbreaking and long-awaited progress on countering a principal threat to the survival of Hawai`i’s unique avifauna.Loss of the Center and its birds would have devastated post-suppression efforts to rebuild and restore bird populations in the wild.
The Post carried a second story about the effort to protect Hawai`i’s endangered birds – a full page of print, even longer – with many photos, on the web. The article mentions the “Birds, Not Mosquitoes” program and varying views about it. I rejoice that the dire situation for the Islands’ biodiversity is getting attention in the Nation’s capital. Again, see my earlier blog.
Plant Invasions in Hawaiian Forests
A team of scientists from the USDA Forest Service and Natural Resources Conservation Service, plus the Hawaii Division of Forestry and Wildlife, has carried out a new assessment of the extent of invasive plant species in forests on the Hawaiian Islands (Potter et al. 2023; full citation at end of blog).
The results of their analysis are – in their words – “sobering”. They portend “a more dire future for Hawai`i`s native forests.”
First, regarding the recent fires, Potter et al. found significantly higher cover by invasive grasses on Forest and Inventory Analysis (FIA) plots on Hawai‘i and Maui than on O‘ahu, Kaua‘i, and Lana‘i. Grass invasions were particularly high on the eastern coast of Maui – near Lahaina. Even so, the authors say their study’s methods resulted in a gross underestimate of areas invaded by fire-prone grasses. That is, most of Hawai’i’s xerophytic dry forests were converted to grasslands before the FIA program began. Therefore these grasslands are not included in FIA surveys.
Psidium cattleyanum; photo by Forrest and Kim Starr
The extent of current invasions in wetter forests is already significant – but trends point to an even more worrying future.
Naturalized non-native plant taxa constitute half of the Hawaiian flora.
56% of Hawaii’s 553,000 ha of forest land contained non-native tree species; about 39% of these forest lands are dominated by non-native tree species. Invasive plant species of particular concern were found in the understory of 27% of surveyed forest plots.
Across all islands, six of the ten most abundant species are non-native: Psidium cattleyanum,Schinus terebinthifolius, Leucaena leucocepahala, Ardisia elliptica, Psidium guajava, and Acacia confusa.
While less than one-third (29%) of large trees across the Islands are non-native, this proportion increases to about two-thirds of saplings (63%) and seedlings (66%). Potter et al. focus on the likelihood that plant succession will result in transformation of these forests’ canopies from native tree species to non-native species.
75% of forests in lower-elevation areas of all islands are already dominated by non-native tree species. “Only” 31% of higher-elevation forests are so dominated. These montane forests have been viewed as refugia for native species, but all are invaded to some extent – and likely to become more degraded.
Potter et al. say the high elevation forests might be more resistant to domination by non-natives. Such a result would be counter to well-documented experience, though. Even the authors report that the montane rainforests and mesophytic forests of O‘ahu and Kaua‘i are heavily invaded by non-native tree species. Such species constitute 86% or more of large trees, saplings, and seedlings in mesophytic forests; 45% of large trees and 66% of seedlings in their montane rainforests.
The most abundant tree species in Hawai`i is the invasive species Psidium cattleyanum (strawberry guava). It was recorded on 88, or37%, of 238 FIA plots. There are nearly twice as many P. cattleyanum saplings as Hawai`i’s most widespread native species, ‘ohi’a lehua (Metrosideros polymorpha).
Widescale replacement of native trees by non-native species is likely. Several factors favor these changes: 1) tree disease – rapid ‘ohi’a death has had drastic impacts on ‘ohi’a populations on several islands; 2) invasions by forbs and grasses; 3) soil damage and other disturbances caused by invasive ungulates; and 4) climate change. If succession conforms to these trends, non-native tree species could eventually constitute 75% or more of the forest tree stems and basal area on all islands and across forest types and elevations.
Loss of Hawai’i’s native tree species would be disastrous for biodiversity at the global level. More than 95% of native Hawaiian tree species are endemic, occurring nowhere else in the world.
The authors analyzed plant presence data from 238 FIA plots. Plots spanned the state’s various climates, soils, elevations, gradients, ownership, and management. However, access issues precluded inclusion of forests from several islands: Moloka‘i, Kaho’olawe, and Ni‘ihau. I know that Moloka‘i, at least, has a protected forest reserve (a Nature Conservancy property) at the island’s highest elevations.
Protecting Native Trees
Federal, state, and private landowners have carried out numerous actions to protect native forests. These efforts might be having some success. For example, forests on public lands, in conservation reserves, or in areas fenced to exclude ungulates were less impacted by non-native plants than unfenced plots, on average. However, the authors could not determine how much of this difference was the result of management or because protections were established in forests with the lowest presence of IAS species. Fencing did not prevent invasions by forbs and grasses – possibly because they are so widespread that seed sources are everywhere.
Hawaii’s two National parks (Hawai`i Volcanoes and Haleakala) have made major efforts to control invasive plants. Hawai`i Volcanoes, on the Big Island, began its efforts in the 1980s; Haleakala (on Maui) more recently. This might be one explanation for the fact that a smaller proportion of the forests on these two islands have been invaded. These efforts have not fully protected the parks, however. Low elevation native rainforests now have a high presence of non-native shrubs. Such forests on Hawai`i Island also have significant invasions by non-native woody vines, forbs and grasses.
More discouraging, intensive efforts have not returned lowland wet forest stands to a native-dominated state. Native tree species are not regenerating—even where there is plentiful seed from native canopy trees and managers have repeatedly removed competing non-native understory plants.
Potter et al. conclude that other approaches will be needed. They suggest deliberate planting of native and non-invasive non-native species or creation of small artificial gaps that might facilitate recovery of native tree species. In montane forests on Hawai`i and Maui, where native tree seedlings account for more than 70% of all tree seedlings, they propose enhancing early detection/rapid response efforts targetting invasive forbs. This would include both National parks.Certainly Haleakala National Park has this priority in mind. It launched a serious effort to try to eradicate Miconia calvescens when this tree first was detected.
Lloyd Loope, much-mourned scientist with US Geological Survey, attacking Miconia on Maui
Potter et al. note the challenge of managing remnant xerophytic dry forests, where natural regeneration of native plants has been strongly limited by invasive grasses; loss of native pollinators and seed dispersers; and the increasing frequency and intensity of droughts. They note that expanded management efforts must be implemented for decades, or longer, to be successful.
Native Trees at Risk to Nonnative Insects
Beyond the scope of the Potter et al. study is the fact that at least two dry forest endemic trees have faced their own threats from non-native insects.
The Erythrina gall wasp, Quadrastichus erythrinae, appeared in Hawai`i in 2005; it originates in east Africa. It attacks the endemic tree, wiliwili, Erythrina sandwicensis. I believe a biocontrol agent, Eurytoma erythrinae, first released in 2008, has effectively protected the wiliwili tree, lessening this threat.
The Myoporum thrips, Klambothrips myopori, from Tasmania, was detected on the Big Island in 2009. It threatens a second native tree. Naio, (Myoporum sandwicense), grows in dry forests, lowlands, upland shrublands, and mesic and wet forest habitats from sea level to 3000 m. The loss of this species would be both a signifcant loss of native biodiversity and a structural loss to native forest habitats. The thrips continues to spread; a decade after the first detection, it was found on the leeward (dry) side of Hawai`i Island with rising levels of infestation and tree dieback.
Rhus sandwicensis on Maui; photo by Forrest and Kim Starr
Two native shrubs, Hawaiian sumac Rhus sandwicensis and Dodonea viscosa, might be at risk from a biocontrol agent in the future. APHIS has approved a biocontrol for the highly invasive Brazilian pepper, Schinus terebinthifolia. Brazilian pepper is the second-most abundant non-native tree species in the State. It was found on 28 of 238 (12%) FIA plots. However, the APHIS-approved biocontrol agent is a thrips—Pseudophilothrips ichini. It is known to attack both of these two native Hawaiian shrubs. The APHIS approval allowed release of the thrips only on the mainland US. However, many insects have been introduced unintentionally from the mainland to Hawai`i. Furthermore, Hawaiian authorities were reported to be considering deliberate introduction of P. ichini to control peppertree on the Islands.
In Conclusion
In conclusion, Potter et al. found that most Hawaiian forests are now hybrid communities of native and non-native species; indeed, a large fraction are novel forests dominated by non-native trees. Business-as-usual management will probably mean that the hybrid forests – and probably those in which the canopy is currently dominated by native species—will follow successional trajectories to novel, non-native- dominated woodlands. This likelihood results in a more dire future for native plants in Hawaiian forests than has been previously described.
Potter at al. hope that their findings can guide research and conservation on other islands, especially those in the Pacific. However, Pacific islands already have the most naturalized species globally for their size—despite what was originally considered their protective geographic isolation.
SOURCE
Potter, K.M., C. Giardina, R.F. Hughes, S. Cordell, O. Kuegler, A. Koch, E. Yuen. 2023. How invaded are Hawaiian forests? Non-native understory tree dominance signals potential canopy replacement. Landsc Ecol https://doi.org/10.1007/s10980-023-01662-6
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Ponderosa pine, Coconico National Forest; photograph by Brady Smith, USFS
Have you noticed, as I have, a spurt of interest in conservation of trees? I can rejoice that more people now focus on this!!!!
I have blogged previously about international and national efforts to determine not only native species deserving conservation priority – by the Morton Arboretum and IUCN but also species most threatened by non-native pests. I have also reported on growing attention to breeding tree resistance to non-native pests.
Some scientists are now focusing on species’ regeneration as a way to understand the probable future of both native and introduced species. I hope that scientists will integrate these new data with existing information on the impacts of invasive non-tree plants and tree-killing introduced pests. We need such a comprehensive picture. That will be a challenge!
Also, I hope attempts to set conservation priorities will influence decisions by governmental and non-governmental funders – and those who influence them! So far, I see little evidence that these key players are paying attention. Some Forest Service scientists and academics are pushing for expanded resistance-breeding efforts. Others are writing sophisticated analyses of non-native pests’ ecosystem impacts. But is the USDA leadership supporting stronger pest-prevention measures? Or funding for research on restoration of species? Are conservation NGOs addressing introduced forest pests?
Here, I summarize new work by Kevin Potter and his colleagues, published in two papers (full references at the end of this blog). After reading my summary, I’d like to know: What do you think? Do you agree with the focus on individual species’ regeneration to set conservation and control priorities? Do you agree with the priority species and geographic regions they suggest?? How should we resolve inconsistencies compared to the priorities suggested by the IUCN and Morton Arboretum? If you do agree, how would you suggest we move forward? If not, what approach do you think would be more useful?
A New Approach to Evaluating Species at Risk
Potter and Riitters (2022) point out that a species’ successful regeneration is key to its population’s future genetic diversity. That, in turn, determines the organisms’ ability to adapt to environmental stress and change. The latter includes, but is not limited to, climate change. Because trees are immobile and long-lived, their populations probably require substantially more genetic variation than those of other kinds of plants.
Potter and colleagues (both articles) used FIA survey data to examine regeneration rates by both tree species native to the continental United States (= CONUS) and non-native tree species introduced to CONUS, Hawai`i, or Puerto Rico. I rejoice that they have included these tropical islands, which are part of the United States and are centers of plant endemism. (Two other blogs provide details on their findings in Hawai`i and Puerto Rico.
Native Trees at Risk: Focus on Poor Regeneration
For CONUS, Potter and Riitters (2022) asked whether 280 native forest tree species are regenerating at sustainable levels, both across their full ranges and in regional portions of their ranges, defined by provisional seed zones (an area within which plant materials are assumed to be adapted). Tree species for which FIA surveys placed 75% of the stems in the sapling or seedling classes are determined to be regenerating at sustainable levels. Tree species exhibiting lower proportions of their stems in these “small tree” classes are said to be failing to regenerate adequately.
Potter and Riitters (2022) found that 46 of the 280 native tree species (16.4%) might be at risk of losing important levels of genetic variation (see the list of species in Table 2 of the article). These included high proportions of species evaluated in the following genera: two of three Platanus species; two of four Nyssa species; about 40% of Juniperus and Pinus; and five of 46 Quercus species (10.9%).
[Many areas of the eastern forest, especially in the Mid-Atlantic region, are reported by Stout, Hille, and Royo (2023) to be have insufficient advance regeneration to replace canopy trees.]
Some species appear to be headed toward outright extinction, not only loss of genetic diversity. These include four relatively rare species in California: Pinus muricata, Platanus racemosa, Pseudotsuga macrocarpa, and Sequioadendron giganteum.No seedlings or saplings are recorded on the plots on which they occurred. I note that Platanus racemosa in southern California is being attacked and killed by the Fusarium dieback vectored by the polygamous and Kuroshio shot hole borers.
Platanus racemosa riddled by invasive shot hole borer; photo by Beatriz Nobua-Behrmann, University of California Cooperative Extension
I find it alarming that a few of the possibly at-risk species have extremely wide distributions. These are Populus deltoides (eastern cottonwood), Platanus occidentalis (American sycamore), and ponderosa pine (Pinus ponderosa). Another group of species are classified as at potential risk in all their seed zones: Juniperus californica, Juniperus osteosperma, Pinus pungens, and Quercus lobata (valley oak). I note that valley oak is also under attack by the recently introduced Mediterranean oak borer. Its vulnerability is exacerbated by its relatively small range.
Potter and Riitters (2022) found distinct geographic hot spots: 15 at-risk species occur primarily in the Southeast and 14 species are in California; both represent nearly a third of the at-risk species.
In general, high rates of regeneration failure are seen in the West. Nine at-risk species (19.6% of the 46) grow in the Southwest, eight in Texas (17.4%), and four in the Rocky Mountains (8.7%). However, the Northeast and Midwest are not immune. Seven species from the former and six from the latter are also regenerating poorly. Considering pines alone, seven of 14 at-risk speciesare in the West and five in the Southeast.
Seed Zones: a Proxy for Local Genotypes
As I noted at the beginning, Potter and Riitters (2022) used USDA Forest Service provisional seed zones as a proxy for areas in which a species is presumably locally adapted. In addition to the 46 species considered failing to regenerate adequately throughout their entire ranges, Potter and Riitters (2022) determined that another 39 species are at potential risk of losing locally adapted genotypes. That is, their regeneration levels fell below the threshold in at least half of the seed zones in which they occurred. These potentially at-risk species are in the same taxonomic groups: 13 pines (33.3% of the 39 species in the category), six junipers (15.3%), and three oaks (7.7 %). These, too are concentrated in the Southeast and California: 40% are in the former — including both bald-cypress species — and 30.8% are in California. Another seven species (17.9% of the 39) are in Texas. The Midwest is home to seven species, the Northeast and Southwest each has five species (12.8%), and the Rocky Mountain region has three species (7.7%).
Bald-cypress; photo by Kej605 via WikiMedia
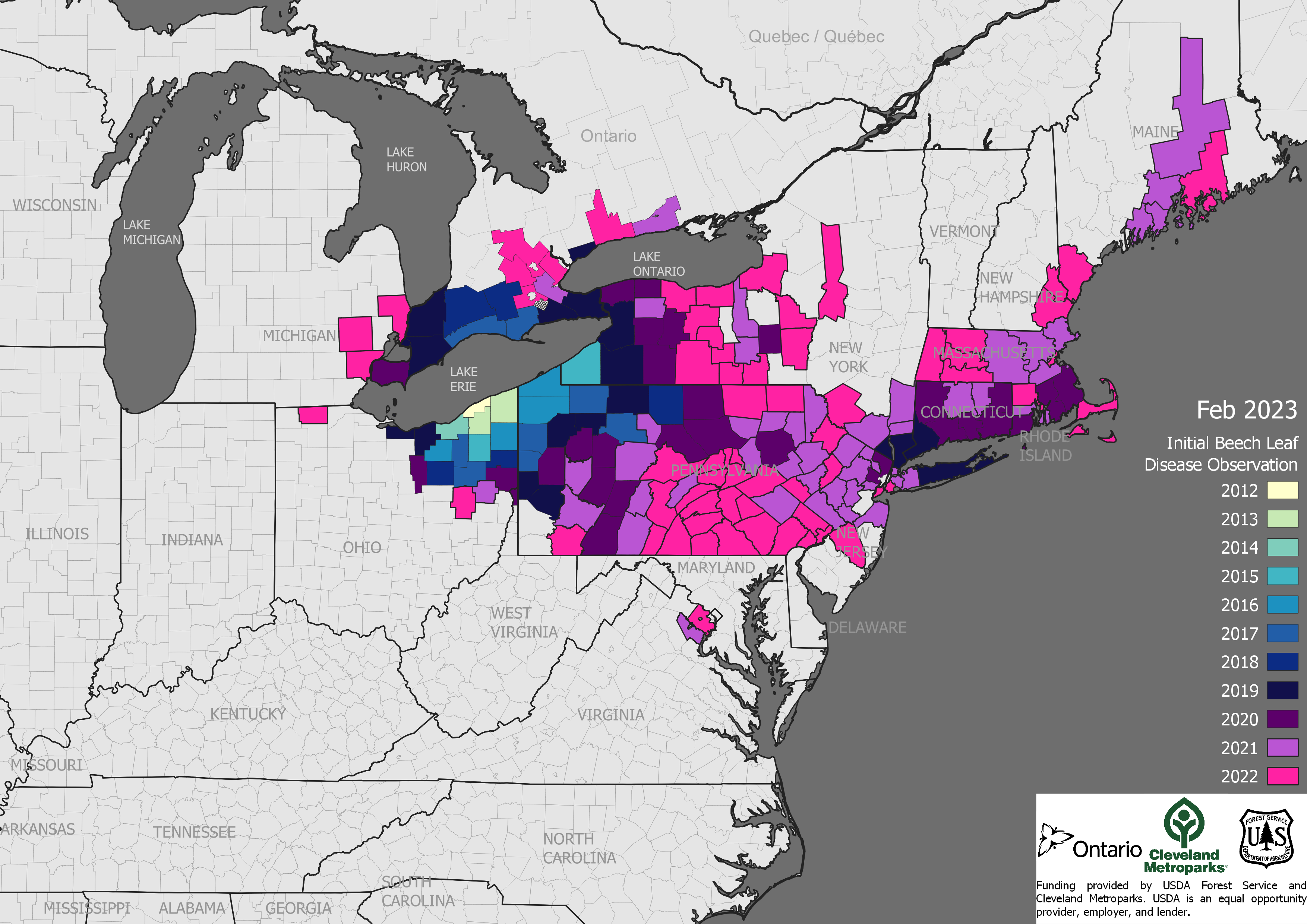
The seed zones with the largest numbers of species regenerating poorly are in the East, specifically the central Great Lakes region, western New York and Pennsylvania, along the Mid-Atlantic and New England coasts, and the coastal plain from southern South Carolina to eastern Texas. Potter and Riitters (2022) say these areas have such high numbers of at-risk species because they are home to so many tree species. I note [although Potter and Riitters (2022) do not] that these regions have also experienced severe levels of tree mortality due to the emerald ash borer (mature and young trees), beech leaf disease (primarily young trees), and laurel wilt disease (sub-canopy trees).
A different geographic pattern appears when considering the proportion — rather than the number — of species facing deficits in regeneration. In several Western regions, 60 – 100% of the tree species fell below the study’s threshold of 75% of recorded stems being in the sapling or seedling sizes. These seed zones are found particularly in parts of California, the Southwest, the Great Basin, and the Pacific Northwest. In none of the seed zones in the East are more than 50% of tree species in the category of potentially losing genetic variation. The implication is that while more species might be lost from parts of the East, the loss of fewer species in some Western seed zones could result in larger impacts on the composition, structure, and function of forest ecosystems there.
Potter and Riitters (2022) say that their approach has limitations because it relies on an assumption that a lack of smaller (i.e., younger) trees is an indication that a species has inadequate regeneration across all or part of its distribution and thus is vulnerable to losing genetic variation. They are not able to quantify directly the genetic variation within most forest tree species. In addition, the choice of 75% or fewer of all trees being seedlings or saplings threshold as the threshold is arbitrary. They believe these decisions are defensible.
Potter and Riitters (2022) hope that indicators of forest sustainability such as this can bridge the gap between scientists, forest managers, policy makers, and other stakeholders.
Further, the authors hope that this approach will help prioritize species most in need of: 1) monitoring for genetic diversity, 2) in situ conservation, and 3) ex situ propagule collections. In a future blog I will compare the species highlighted by Potter and Riitters (2022) to the earlier priority list developed by the IUCN and Morton Arboretum. Finally, the focus on regeneration levels could help scientists design representative sampling protocols for range-wide ex situ propagule collections for genetic diversity studies using molecular markers.
Applying This Analysis to Invasions by Non-native Trees
In a second study, Potter, Riitters, and Guo (full citation at end of this blog) flipped the focus: they used the same approach to quantify the degree of invasion by non-native trees in the U.S. I’ve blogged about this study, in general, here. Also see my separate blogs for its welcome application to Hawai`i and Puerto Rico.
Again, Potter, Riitters, and Guo hope their approach will assist in the crucial, difficult task of distinguishing between high-impact and less threatening non-native species. They warn, however, that the FIA survey procotol does not suit the needs of an early detection system.
Differentiating Invasive Tree Species’ Impacts
Potter, Riitters, and Guo note that thousands of non-native tree species have been planted around world to provide an extensive list of ecosystem services. Globally, 400 tree species have been recognized as naturalized (= consistently reproducing) or invasive (= spreading) in areas outside their native ranges. Contrary to some expectations, even relatively undisturbed forests are affected by invasive plants. In the continental United States, many fewer invasive plant species are trees than other forms/habits – shrubs, forbs, gramminoids. On the tropical islands, a much higher proportion of invasive plants are trees.
Lugo et al. (2022; full citation at end of this blog) find non-native tree species occupy a tiny fraction of the forest area of the continental United States [= CONUS], i.e., only 2.8% of the area, and only 0.4% of all tree species recorded in the FIA plots. However, these non-native tree species are widespread. They are found in 61% of forested ecosections in CONUS. Also, they are becoming more common in invaded sites. [Ecosections are divisions within 37 ecological provinces in the hierarchical framework developed by Cleland et al. (2007). There are 190 ecosections in U.S. forest biomes.]
Potter, Riitters, and Guo categorized those non-native tree species with at least 75% of stems detected by FIA surveys to be in sapling or seedling size as highly invasive. In other words, these species are successfully reproducing after reaching the canopy. So they might be more likely to alter forest functions and ecosystem services than those reproducing less robustly. They classified as species with 60 – 75% of recorded stems in these “small tree” categories as “moderately invasive.”
Potter, Riitters, and Guo suggest that control might more productively target the moderately invasive species in geographic regions where they have spread less so far – so presumably fewer seed-bearing mature specimens are present. They list as examples Picea abies, Pinus sylvestris, and Paulownia tomentosa.
In CONUS, FIA protocols specify reporting of 30 non-indigenous tree species.
Acer platanoides
Ailanthus altissima
Albizia julibrissin
Alnus glutinosa
Castanea mollissima
Casuarina lepidophloia
Cinnamomum camphora
Citrus sp.
Elaeagnus angustifolia
Eucalyptus globulus
Eucalyptus grandis
Ginko biloba
Melaleuca quinquenervia
Melia azedarach
Morus alba
Paulownia tomentosa
Picea abies
Pinus nigra
Pinus sylvestris
Populus alba
Prunus avium
Prunus persica
Salix alba
Salix sepulcralis
Sorbus aucuparia
Tamarix spp
Triadica sebifera
Ulmus pumila
Vernicia fordii
About half of these –16 species – qualified under the Potter, Riitters, and Guo criteria as highly invasive: Acer platanoides, Ailanthus altissima,Albizia julibrissin, Cinnamomum camphora, Elaegnus angustifolia, Melia azedarach, Melaleuca quinquenervia, Morus alba, Picea abies, Pinus nigra, Prunus avium, Salix alba, Salix sepulcralis, Triadica sebifera, Ulmus pumila, Vernicia fordii. An additional four taxa are ranked as potentially highly invasive: Tamarix; Eucalyptus grandis and E. globulus, Populus alba.
ring-billed gulls eating berries of Chinese tallowtree (Triadica sebifera); photo by TexasEagle via Flickr
I ask : Do YOU agree that these taxa are the most important to be tracking as potentially invasive in forests of the continental United States?
Potter, Riitters, and Guo distinguish between the most “common” and the most “widespread” invasive tree species – although they do not define the differences. Some of the most “common” or “widespread” species are not a surprise: Ailanthus altissima, Triadica sebifera (syn. Sapium sebiferum), and Acer platanoides. Ailanthus is categorized as highly invasive in 39 of 44 ecoregions in which it occurs. It is also notoriously difficult to manage. Triadica sebifera is classified as highly invasive in every one of the 20 ecoregions in which it occurs. It produces prolific seed crops that are widely dispersed by birds and water. It can invade both disturbed and undisturbed habitats. Some of the common or widespread species do surprise me: Ulmus pumila, Morus alba and Picea abies.
Most of the non-native tree species occur on only 2% of plots in the ecoregions in which they occur. However, some highly invasive trees exceed this level:
Triadica sebifera is detected on 8.6% of plots on average across 20 ecoregions;
Ulmus pumila is detected on 3.7% of plots across 39 ecoregions;
Elaeagnus angustifolia is detected on 3.3% of plots in 13 ecoregions;
Melaleuca quinquenervia is detected on 2.7% of plots in 4 ecoregions.
A. altissima is detected on only 2% of plots in the 44 ecoregions. This is surprising to me. I see it everywhere in the Mid-Atlantic – and elsewhere!
[In USFS Region 9 (24 states in the Northeast and Midwest), FIA surveys in 2019 detected Ailanthus on only 3% of plots, Norway maple and Siberian elm each on only 1% of plots (Kurz 2023).]
Eastern U.S. forests are invaded at rates several times those in Western forests, both as a proportion of plots that are invaded and the diversity of plant growth forms. The probability of invasion is highest in Eastern forests that are relatively productive and located in fragmented landscapes that contain developed or agricultural land. Non-native invasive trees are most prevalent along the Gulf Coast and in Mid-Atlantic and Midwestern States. Highly invasive non-native trees are most diverse in the ecoregions of the Mid-Atlantic and Southeast. I note that these regions also rank high in numbers of native tree species determined by Potter et al.’s other study to be reproducing an unsustainable levels.
The study found that non-native trees are almost entirely absent from the Rocky Mountain States and Alaska. However, I have seen Ailanthus in riparian areas of Utah, Arizona, and New Mexico. While few non-native tree species are recorded from ecoregions along the Pacific Coast, those areas are heavily invaded by other types of plants. Lugo et al. say those shrubs and forbs are not interfering with forest regeneration. Do YOU agree?
BLM & USFS botanists removing Spanish broom from Rogue River Canyon; photo by Stacy Johnson, BLM
On tropical islands included in the study – Hawai`i and Puerto Rico – the situation is very different. Together, these islands’ tree canopy covers less than 0.5% that of the area in the lower 48. Hawai`i is recognized as a global hotspot of non-native species richness. Naturalized non-native plant taxa constitute about half of the Hawaiian flora. The US Forest Service tracks twice as many non-native tree species in Hawai`i (62) than over the entire continental U.S. plus Alaska.
Of these 62 species, Potter, Riitters, and Guo identified 26 tree species as either highly or moderately invasive, either already or potentially highly invasive, three as moderately invasive, seven as potentially moderately invasive. In general, the richness of non-native tree species is higher in lower-elevation ecoregions, especially the lowland/leeward dry and mesic forests on O’ahu and lowland wet and mesic forests of the Big Island. [The article makes a brief reference to the probable role of rapid ʻōhiʻa death opening the canopy of the mesic and wet forests, thereby facilitating plant invasions.]Most Hawaiian ecoregions, especially those on O’ahu and Hawai’i Island, had higher non-native tree species richness than even the most highly invaded ecoregions in the lower 48 states. Parts of O’ahu & Maui had the most non-native tree species classified as highly invasive.
The Caribbean archipelago – including but not limited to Puerto Rico – has a lower proportion of non-native plant species than Hawai’i — 17% of plant species are not native. However, their presence is even higher: two-thirds of Puerto Rico’s forests comprise novel tree assemblages. This is probably because Puerto Rico has half the land area of the Hawaiian archipelago and has been part of global trade networks for 500 years instead of 200. Potter and colleagues identified 17 non-native tree species as highly invasive, 16 as potentially highly invasive, and two as moderately invasive.
On the continent only seven of 30 non-native tree species occurr on at least 2% of FIA plots across the ecoregions in which they are inventoried. Hawai’i is stunningly different: 56 of 62 species occurr on at least 2% of plots across ecoregions on average; 24 species are present on at least 10% of plots on average. One species, Psidium cattleyanum, is present on nearly half of surveyed plots across 13 ecoregions! In Puerto Rico, 21 species occurred on at least 2% of the FIA plots.
Acacia confusa – highly invasive in dry forests of Hawai`i; photo by Forrest and Kim Starr
Potter, Riitters, and Guo could not assess the invasiveness of several species that occurred only as small stems in a couple of plots. There are 11 such species on Hawai`i, eight on Puerto Rico. These species might be in the early stages of widespread invasion, or they might never be able to reproduce and spread. Despite the uncertainty, the authors suggest that eradication or control efforts targetting these species might be more cost-effective since presumably few trees have reached reproductive age yet. In Puerto Rico, they single out Schinus terebinthifolius, since it is already recognized as moderately invasive in Hawai`i [I add – seriously invasive in nearby Florida!]. However, they also emphasize the threat from one of the widespread species, Syzgium jambos, because it is a shade-tolerant species that can form dense, monotypic stands under closed canopies
I have posted separate blogs providing more details on the invasive tree species in Hawai`i and Puerto Rico.
Limits of the FIA Dataset
As in the study of native species regeneration, Potter, Riitters, and Guo specify limits arising from use of the FIA dataset. Two seem particularly pertinent to evaluation of the situation on the tropical islands.
First, they cataloged only those non-native tree species chosen by the FIA program administrators to track in the three major regions. Again, I ask YOU whether you agree with the species being recorded. Should others species be included? Should some of these species be dropped?
Second, the survey protocol does not differentiate between sites with significantly different status and history. For example, non-native trees growing on abandoned agricultural sites are counted the same way as those growing in presumably old-growth forests. They conclude that including such sites might explain the records of Eucalyptus and pine species in surveys on the islands.
Finally, as noted in the other study, the program incorporates plots that contain at least 10% canopy cover by live trees or had such cover in the past. The inventory has not included urban parks – although in recent years an urban inventory protocol has been developed.
I remind you that Potter, Riitters, and Guo warned that the FIA inventory is not designed to detect newly introduced species that are early in the invasion process.
SOURCES
Kurtz, C.M. 2023. An assessment of invasive plant species in northern U.S. forests. Res. Note NRS-311. http://doi.org/10.2737/NRS-RN-311
Lugo, A.E., J.E. Smith, K.M. Potter, H. Marcano Vega, and C.M. Kurtz. 2022. The Contribution of NIS Tree Species to the Structure and Composition of Forests in the Conterminous United States in Comparison with Tropical Islands in the Pacific & Caribbean. USDA USFS General Technical Report IITF-54.
Potter, K.M and Riitters, K. 2022. A National Multi-Scale Assessment of Regeneration Deficit as an Indicator of Potential Risk of Forest Genetic Variation Loss. Forests 2022, 13, 19. https://doi.org/10.3390/f13010019.
Potter K.M., Riitters, K.H. and Guo, Q. 2022. Non-native tree regeneration indicates regional and national risks from current invasions. Frontiers in Forests and Global Change doi: 10.3389/ffgc.2022.966407
Stout, S.L., A.T. Hille, and A.A. Royo. 2023. Science-Management Collaboration is Essential to Address Current & Future Forestry Challenges. IN United States Department of Agriculture. Forest Service. 2023. Proceedings of the First Biennial Northern Hardwood Conference 2021: Bridging Science and Management for the Future. Northern Research Station General Technical Report NRS-P-211 May 2023
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
white ash: a species that might be restored under the programs envisioned in the proposed bills
Bills have been introduced into both the House and Senate to enhance USDA APHIS and Forest Service programs intended to curtail introduction and spread of non-native forest pests and disease and – especially – programs aimed at restoring pest-decimated trees to the forest.
The House bill is H.R. 3174; it was introduced by Reps. Becca Balint (VT).
The Senate bill is S. 1238; it was introduced by Senators Peter Welch (VT), Mike Braun (IN), and Maggie Hassen (NH). [Both senators Welch and Braun are on the Agriculture Committee – which will write the bill.]
CISP hopes that the contents of these two bills will be incorporated in the Farm Bill that Congress is expected to adopt this year or next. The proposals have the support of the Forests in the Farm Bill coalition. [Unfortunately, neither the “Consolidated Recommendations” nor “Summarized Recommendations appears to be posted on the internet at present.]
In the last Congress, a nearly identical bill introduced by then-Representative Peter Welch was endorsed by the organizations listed below. We hope they will endorse the new bills now! If you are a member of one of these organizations, please ask them to do so.
Organizations that endorsed the previous bill: Vermont Woodlands Association, American Forest Foundation, Center for Invasive Species Prevention, Reduce Risk from Invasive Species Coalition, National Woodland Owners Association (NWOA), National Association of State Foresters (NASF), The Society of American Foresters (SAF), the North American Invasive Species Management Association (NAISMA), the Ecological Society of America, Entomological Society of America, a broad group of university professors and scientists, The Nature Conservancy (TNC) Vermont, Audubon Vermont, the Massachusetts Forest Alliance, the New Hampshire Timberland Owners Association, the Maine Woodland Owners Association, and the Pennsylvania Forestry Association.
I seek your help in generating support for incorporating these proposals into the 2023 Farm Bill. Please urge your representative and senators to co-sponsor the bills or otherwise support that action.
beech in a breeding experiment at The Holden Arboretum; photo by Jennifer Koch
Key points of the two bills:
They strengthen APHIS’ access to emergency funds. APHIS has had the authority to access emergency funds from the Commodity Credit Corporation since 2000. However, the Office of Management and Budget has often blocked its requests. See § 2, of the bills, EMERGENCY AUTHORITY WITH RESPECT TO INVASIVE SPECIES.
It creates two separate but related grant programs.
The first grant program – in § 3. FOREST RECLAMATION GRANTS – funds research addressing specific questions impeding the recovery of tree species that are native to the US and have suffered severe levels of mortality caused by non-native plant pests or noxious weeds.
The second grant program – in § 4. FOREST RESTORATION IMPLEMENTATION GRANTS – funds implementation of projects to restore these pest-decimated tree species to the forest. These projects must be part of a forest restoration strategy that incorporates a majority of the following components:
(1) Collection and conservation of native tree genetic material.
(2) Production of propagules of the target tree species in numbers sufficient for landscape-scale restoration.
(3) Preparation of planting sites in the target tree species’ former habitats.
(4) Planting of native tree seedlings.
(5) Post-planting maintenance of native trees.
§ 5 states that the absence of a national policy on addressing nonnative forest pests has resulted in their receiving a low priority within all Federal agencies. It then mandates a study to analyze agencies’ available resources, raise the issue’s priority, and improve coordination among agencies. This study is to be carried out by an independent institution, for example the National Academy of Sciences. The authors are to consult with specialists in entomology, genetics, forest pathology, tree breeding, forest and urban ecology, and invasive species management.
Funding for all three action components – the emergency response and both grant programs – would come from the Commodity Credit Corporation, so it would not be subject to the vagaries of annual appropriations bills.
Forest Restoration Alliance volunteers potting hemlock seedlings; photo provided by Fred Hains
Entities which could apply for the research grants (§ 3 of the bills) include Federal agencies; State cooperative institutions; academic institutions offering degrees in the study of food, forestry, and agricultural sciences; and non-profit organizations exempt from taxes under §501(c)(3) of the tax code. Types of research funded could include:
‘‘(A) biocontrol of nonnative pests & diseases or noxious weeds severely damaging native tree species [the bill does not specify, but Project CAPTURE identifies many qualifying species; see also my earlier blog];
‘‘(B) exploration of genetic manipulation of the plant pests or noxious weeds;
‘‘(C) enhancement of pest-resistance mechanisms of hosts; and
‘‘(D) development of other strategies for restoring individual tree species.
The maximum amount of such grants is $400,000 per year.
Entities which could apply for the implementation grants (§ 4 of the bills) include a cooperating forestry school; a land-grant college or university; a State agricultural experimental station; a 501(c)(3) organization. Funding would begin at $3 million for FY 2023 and rise to $10 million for FY 2026.
The Secretary of Agriculture would be guided in implementing these programs by two committees. One – the committee of experts – would constitute representatives of the USFS, APHIS, ARS & State forestry agencies. The second – the advisory committee – would be composed of representatives of land-grant colleges and universities and affiliated State agriculture experiment stations, forest products industry, recreationists, and professional forester, conservation, and conservation scientist organizations.
Port-Orford cedar seedlings at USFS Dorena Center – a model for success! Photo provided by Richard Sniezko
Please contact your Member of Congress (Representative) and senators to urge them to support inclusion of these provisions in the Farm Bill. [Remember: they work for us!] Telling them of your support for these bills is especially important if your Representative or Senator is on the Agriculture Committee. I list those legislators here:
State
HOUSE AGRIC COMM
SENATE AGRIC COMM
AL
Barry Moore
Tommy Tuberville
AR
Rick Crawford
John Boozman
CA
Doug Lamalfa John Duarte Jim Costa Salud Carbajal
CO
Yadira Caraveo
Michael Bennet
CT
Jahana Hayes
FL
Kat Cammack Darren Soto
GA
Austin Scott David Scott Sanford Bishop
Raphael Warnock
HI
Jill Tokuda
IA
Randy Feenstra Zach Nunn
Joni Ernst Charles Grassley
IL
Mike Bost Mary Miller Nikki Budzinski Eric Sorensen Jonathan Jackson
Richard Durbin
IN
Jim Baird
Mike Braun
KS
Tracey Mann Sharice Davids
Roger Marshall
KY
Mitch McConnell
MA
Jim McGovern
ME
Chellie Pingree
MI
Elissa Slotkin
Debbie Stabenow
MN
Angie Craig
Amy Klobuchar Tina Smith
MO
Mark Alford
MS
Trent Kelly
Cindy Hyde-Smith
NC
David Rouzer Alma Adams
ND
John Hoeven
NE
Don Bacon
Deb Fischer
NJ
Cory Booker
NM
Gabe Vasquez
Ben Ray Lujan
NY
Marc Molinaro Nick Langworthy
Kirsten Gillibrand
OH
Max Miller Shontel Brown
Sherrod Brown
OK
Frank Lucas
OR
Lori Chavez-Deremer Andrea Salinas
PA
Glenn Thompson
John Fetterman
SD
Dusty Johnson
John Thune
TN
Scott Desjarlais Brad Finstad
TX
Ronny Jackson Monica de la Cruz Jasmine Crockett
VA
Abigail Spanberger
VT
Peter Welch
WA
Marie Gluesenkamp Perez
WI
Derrick van Orden
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
red spruce (Picea rubens) — the conifer at greatest risk; This grove is in Great Smoky Mountains National Park; photo by Famartin via Wikimedia Commons
Scientists have incorporated into the widely-used urban tree management tool, i-Tree, a tool to help predict the damage that an insect species little known in North America might cause to trees growing in a specific area if it is introduced. This tool is available to all here.
I rejoice that predictive tools are becoming widely available. The tool is obviously the result of a lot of work by participating scientists – who are listed below. I hope many of you will try it out! Perhaps you and your students can join efforts by the tool-development team, especially in analyzing insect species from Central America and Asia that have not yet arrived in North America? If you are interested in helping, contact Katheryn Thomas, Angela Mech, or Ashley Schulz; you can obtain their contact information by visiting their institution’s website. You might choose which insect species to evaluate by consulting your own or colleagues’ research, reviewing the refereed and grey literature, APHIS and CFIA interception databases, databases maintained by several countries, websites such as CABI, EPPO, etc.
The new tool might help create a more effective “early warning” system. Whether this happens depends on what others do now. Anyone – perhaps a staffer of a federal or state agency, or a city tree manager, or an academic – can apply the tool to meet his/her own objectives. If a more effective national or continental “early warning” system is to be created, someone needs to set up a process for conveying the findings to responsible federal or state/provincial agencies or even the scientific societies, e.g., Entomological Society (and, in the case of beetles transporting associated fungi, American Phytopathological Society). Perhaps the most challenging issue is to find an entity willing to receive these communications, review their accuracy, and – at a minimum – make the results accessible to phytosanitary agencies, interested public, etc. One possible entity is “PestLens”, a web-based early-warning system maintained by APHIS. The project’s objective is to provide early-warning information and facilitate a prompt, coordinated, and appropriate safeguarding response. PestLens posts alerts once a month. These are visible to anyone who subscribes. However, it remains unclear how often APHIS and state agencies act on the notices. The North American Plant Protection Organization (NAPPO) also hosts an alert system, but it records only official notices, leading to some absurdities. (E.g., NAPPO reported Mexico’s designation of the invasive shot hole borers as quarantine pests – without mentioning that they are well-established in California because neither APHIS nor California Department of Food and Agriculture has designated the insects as officially regulated.)
Those applying the tool need to have some knowledge and access to a range of scientific resources (including, in my view, people who can check the accuracy of the data entered into the system). Users must have appropriate skills to conduct some research into the insect and what it feeds on. Information required for the tool includes the following:
taxonomic information for the insect (Order, Family, Genus, Species)
the feeding guild of the insect (i.e., foliovore, gall, reproductive, root, sap, wood)
climate in the native range of the insect (i.e., Tropical, Dry, Temperate, Continental, Polar)
native range of the insect (i.e., Afrotropical, Australasian, Indomalayan, Neotropical, Oceanian, Palearctic Asia, Palearctic Europe)
the host trees of the insect in its native range (scientific name [Genus species]). The tool warns participants to include the full range of potential tree hosts – by listing either all or a representative sample. The tool will use this information to estimate the evolutionary distance between known native hosts and potential North American hosts using comprehensive phylogenetic tree of plants.
Clearly, those using the tool have their work cut out for them! The tool does provide definitions, descriptors, and drop-down lists for most of the factors, including insect orders and families, tree genera, geographic origins, and climate types. Users are now anticipated to be employees of federal and presumably state agencies; academics – even students!—and others who have the capacity to research what an insect feeds on in its native range.
This tool is intended to predict the probability that an insect species of concern – either newly detected in the country or thought likely to invade based on port detections or other reasons — will become a high impact invader. I rejoice that they are inclusive – the tool can test the vulnerability of 50+ conifer species and 360+ hardwood species native to North America. Assuming the assessor can enter accurate information for the categories outlined above, the tool can then provide a list of probabilities for each relevant North American host tree.
The tool is based on the findings of two studies, Mechet al. and Schulzet al. (full citations at the end of this blog). I discussed these studies in earlier blogs. They were also incorporated into the broader effort to identify predictive traits carried out by Raffa et al. (full citations at the end of this blog) and discussed in a separate blog. See the section titled “Potential” to see the exciting results of an application of the Mech et al. findings and methods.
To develop the tool, project scientists synthesized data on traits and factors representing four types of drivers: (1) insect traits, (2) tree traits (especially those associated with host defenses), (3) the relatedness between the insect’s native and North American tree hosts, and (4) the relatedness between the non-native insect and North American insects on the same tree. They tested key hypotheses, e.g., defense free space and enemy release. The team tested the tool with researchers from USDA APHIS and Canadian Food Inspection Agency (CFIA), Northeast Plant Diagnostic Network, and National Invasive Species Council.
Norway spruce (Picea abies) — host of 30 of the 62 insect species analyzed in Uden et al.; photo by Marzena via Pixabay
The research group hopes this tool will stimulate development of a global database of insects which will utilize the results of basic research on phytophagous insects and what they eat. Basic research on insects native to North America is also important and can benefit other countries that might want to develop a similar tool for their own phytosanitary needs.
The Tool’s Potential
Many of the scientists who developed the i-Tree tool have participated in an analysis of the threat to North American conifer species posed by insects native to Europe that have not yet been introduced to North America (Uden et al.). They applied the methodology from Mech et al., which is comparable to, although not identical to, the i-Tree system. They (1) created a list of 62 European insect species that appear to pose a risk to 47 species of North American conifers; (2) identified and compared the predicted likelihoods of high-impact invasion under each of four phylogenetic systems datasets; and (3) evaluated risk and vulnerability trends among insects & conifer hosts, respectively. In total they evaluated 2,914 insect–novel host pairs.
Fraser fir (Abies fraseri) in Great Smoky Mountains National Park; photo by James St. John via Flickr
Among their findings are the following:
Of the 2,914 pairs examined, 302 (10.4%) had a predicted risk of high impact. These pairs included 41 (66%) of the insect species and 20 (41.7%) of the conifer species. The proportion of potential invasions posing a significant risk is higher than those indicated by earlier studies.
The insect species posing a risk of high-impact invasion were spread among insect orders, with relatively high levels concentrated in Lepidoptera and Coleoptera, fewer in the Hymenoptera and Hemiptera.
Consistent with Mech et al., they found a “Goldilocks” period of evolutionary divergence of hosts exposing the North American tree species to the highest risk. Thus, if a North American conifer shared a common ancestor with the insect’s native European host ~2–10 million years ago, it was predicted to be more vulnerable to a high-impact invasion by a conifer specialist.
North American fir (Abies) and spruce (Picea) species are more vulnerable to the introduction of European conifer-specialist insects than are pines (Pinus). [Mech et al. found that trees with high shade tolerance and low drought tolerance are more vulnerable. These traits also fit fir and spruce; but not pine.] The most vulnerable tree species was red spruce (Picea rubens).
Uden et al. also say Fraser fir (Abies fraseri) and Carolina hemlock (Tsuga caroliniana) are highly vulnerable to European insect species. They identified 17 high-risk insect species for Fraser fir. Of course, both are already severely depleted by non-native insect pests (Balsam woolly adelgid and hemlock woolly adelgid, respectively). They have also been identified by the Potteret al. “Project CAPTURE” process as having high priorities for conservation efforts.
I worry that fir and spruce are less important as timber species than pines; I hope this does not result in agencies and important stakeholders assigning this risk finding a lower priority.
Uden et al. assert that their study shows that this system can identify vulnerable tree species in the absence of information about which particular insect might invade. This information helps managers focus biosecurity and management program programs on protecting the most vulnerable tree species. However, 57% of the North American conifers (27 species) were found to be vulnerable under at least one of the insect-host pairs. To further set priorities, they suggest combining predictions from this analysis with USFS Forest Inventory and Analysis (FIA) data to identify vulnerable biogeographic regions and vegetation communities. (Fraser fir and Carolina hemlock rank high under this process.) Scientists could also apply species importance indicators, such as the NatureServe Explorer plant community descriptions. They suggest linking these criteria to the USFS Early Detection Rapid Response surveillance program, link to website which currently targets specific insect species.
red pine (Pinus resinosa) – the pine species at greatest risk; photo by Charles Dawley via Flickr
Uden et al. also warn that their analysis focused on a narrow range of possible introduced species: insects from Europe that feed on conifers exclusively. They caution that no one should assume that tree species that have a low “vulnerability” rank in this study should be considered at low risk for all possible introduced insects. They suggest researchers should identify tree species from the wider Palearctic that are within the high-impact “Goldilocks” zone of divergence times in relation to specific North American tree species, and then identify the insects that feed on those Palearctic trees to determine the species that would have the highest predicted risk of causing a high impact on those North American conifers.
Of course, many North American tree species are not conifers! Applying the methods in Schulz et al. – now integrated into the i-Tree tool – would facilitate similar predictive findings for the angiosperms.
Participants
The importance of this project is seen in the impressive array of funders supporting it. They include:
U.S. Geological Survey John Wesley Powell Center for Analysis and Synthesis for a working group titled “Predicting the nest high-impact insect invasion: Elucidating traits and factors determining the risk of introduced herbivorous insects on North American native plants;”
USDA Forest Service National Urban and Community Forestry Advisory Council funded a working group titled “Forecasting high-impact insect invasions by integrating probability models with i-Tree from urban to continental scales”;
Nebraska Cooperative Fish and Wildlife Research Unit;
University of Washington;
USDA Forest Service Eastern Forest Environmental Threat Assessment;
National Science Foundation Long-Term Ecological Research program;
USDA Forest Service International Programs; and
USDA National Institute of Food and Agriculture (Hatch and McIntire-Stennis projects).
Scientists who created this tool: Kathryn A. Thomas (USGS – Southwest Biological Research Center) Travis D. Marsico (Arkansas State University) Daniel A. Herms (The Davey Tree Expert Company) Patrick C. Tobin (University of Washington) Andrew Liebhold (U.S. Forest Service) Nathan Havill (U.S. Forest Service) Angela Mech (University of Maine) Ashley Schulz (Mississippi State University) Matthew Ayres (Dartmouth College) Kamal Gandhi (University of Georgia) Ruth A. Hufbauer (Colorado State University)
Kenneth Raffa (University of Wisconsin) Daniel
Uden (University of Nebraska-Lincoln)
Carissa Aoki (Maryland Institute College of Art)
Scott Maco (The Davey Tree Expert Company)
Angela Hoover (University of Arizona)
SOURCES
Mech, A.M., K.A. Thomas, T.D. Marsico, D.A. Herms, C.R. Allen, M.P. Ayres, K.J.K Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, and P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecol Evol. 2019. Nov; 9(21):12216-12230.
Potter, K.M., Escanferla, M.E., Jetton, R.M., Man, G., Crane, B.S. 2019. Prioritizing the conservation needs of United States tree species: Evaluating vulnerability to forest insect and disease threats. Global Ecology and Conservation (2019), doi: https://doi.org/10.1016/j.gecco.2019.e00622.
Raffa, K.F., E.G. Brockerhoff, J-C Gregoire, R.C. Hamelin, A.M. Liebhold, A. Santini, R.C. Venette, and M.J. Wingfield. 2023. Approaches to Forecasting Damage by Invasive Forest P&P: A Cross-Assessment. BioScience Vol. 73 No. 2: 85–111 https://doi.org/10.1093/biosci/biac108
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions.
Uden, D.R, A.M. Mech, N.P. Havill, A.N. Schulz, M.P. Ayres, D.A. Herms, A.M. Hoover, K.J. K. Gandhi, R.A. Hufbauer, A.M. Liebhold, T.D. M., K.F. Raffa, K.A. Thomas, P.C. Tobin, C.R. Allen. 2023. Phylogenetic risk assessment is robust for forecasting the impact of European insects on North American conifers. Ecological Applications. 2023; 33:e2761.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
red deer on farm in New Zealand; photo by Bernard Spragg via Flickr
Among the non-native species damaging forest systems are mammals – introduced deer, goats and sheep, and swine, … These animals have the greatest impacts on island systems that are sufficiently isolated that they have no native terrestrial mammals, e.g., Hawai`i and New Zealand. Several New Zealanders have published a study of their impacts (Allen et al.; full citation at end of the blog). The focus of their analysis is the native forests’ ability to sequester carbon and thus mitigate climate change. The scientists are well aware, however, that forests provide many other ecosystem values and services, including biodiversity, water supply and quality, etc.
Introduced ungulates can have many direct effects: reduction and damage to understory biomass, depletion of seedling regeneration, exacerbated soil erosion, and local nutrient imbalances. Mammals’ browsing can modify the composition of plant communities by favoring abundance of unpalatable species. Changes also can alter ecosystem functions associated with nutrient cycling, e.g., by reducing nutrient returns to the soil and altering rates of litter decomposition
In these ways, introduced ungulates exert long-term impacts on forests’ capacity to store carbon.
Allen et al. aimed to determine the extent of these effects on forests’ capacity to store carbon, both above- and below-ground, and on forest structure and diversity. The authors compared data from 26 pairs of sites across New Zealand – half with ungulate exclosures and half adjacent unfenced control plots. The ungulate exclosures had all been established for at least 20 years. All the sites were in species-rich communities of conifers and broadleaved evergreen angiosperm trees. These forests (1) cover about one-third of the country’s remaining mature natural forest; (2) contain tree species of a wide range of palatability to ungulate herbivores; and (3) have been named a conservation priority for forest carbon management. The ungulates present on the plots were European red deer (Cervus elaphus), fallow deer (Dama dama), sika deer (Cervus nippon), and feral goats (Capra hircus).
They assert that New Zealand is a good place to do this type of study because ungulate introductions are relatively recent so their impacts are well documented.
Allen et al. found that managing invasive ungulates makes valuable contributions to conserving biodiversity but not to carbon sequestration. They found little difference in total ecosystem carbon between ungulate exclosures and unfenced control plots. Most of the difference they did find was explained by the biomass of the largest tree within each plot. As they point out, these large trees have been unaffected by invasive ungulates introduced during the last 20–50 years. However, they believe ungulate-caused changes in understory biomass, species composition, and functional diversity might result in major shifts in the diversity and composition of regenerating species. Hence, longer term consequences for both ecosystem processes and storage of forest carbon storage can be expected.
Indeed, excluding ungulates did increase the abundance and diversity of saplings and small trees. The basal area of the smallest class of tree size was 70% greater. Species richness of small trees and saplings was 44% and 68% higher, respectively. This difference had little impact on overall carbon storage, however, because the small trees and saplings store only about 5%. In contrast, the largest tree size class (dbh =/>30 cm), with their roots, contributed 44% of total ecosystem carbon in both exclosure and control plots. The largest effects of exclosures on carbon stocks were in early successional stands, e.g., those affected by such major disturbances as windthrow, volcanic activity, or landslides.
Climate change is expected to cause surprising interactions among forest productivity, herbivory, disturbance. Allen et al. suggest that authorities should focus on excluding ungulates on these highly productive regenerating forests rather than old-growth forests. I am disturbed by this suggestion. It exposes the most biologically diverse forests to continuing damage.
Data gaps
New Zealand has many long-lived, slow-growing tree species. Recruitment of understory trees is already low across both main islands. This situation has been attributed to ungulate browsing. Over centuries, this might result in shifts in the canopy composition. Allen et al. call for additional research to increase our understanding of how browsing and other short-and long-term drivers affect the regeneration of large trees. Also, data on soil CO2 emissions needs better integration.
Australian brushtail possum; photo by Peter Firminger via Flickr
The study did not consider the impact of other introduced mammals, such as feral pigs (Sus scrofa), rodents, and Australian brushtail possum (Trichosurus vulpecula). The possum is known to damage New Zealand trees. The scientists did not explain this omission; I assume it might have been the result of either lack of resources to support a broader study or differences in management strategies – or both?
I note that the study also did not address the extent to which non-native pathogens threaten these large trees. In response to my query, Kara Allen said that their plots did not include many kauri (Agathis australis) trees, so the severe dieback disease caused by Phytophthora agathidicida did not affect their results. Naturally regenerating kauri is limited to a small area of warm temperate rainforests located at the top of the North Island. So kauri potentially play a relatively small role in terms of overall carbon stocks in New Zealand’s forests. On the other hand, Allen says thatmyrtle rust (Austropuccinia psidii) could have a major impact on New Zealand forests’ carbon storage. Trees in the host family, Myrtaceae, are ecologically important across both islands. Also, they comprise a large portion of overall forest carbon stocks (ranked in the top 5 largest families for above- and belowground biomass). An example is southern rata (Mterosideros umbellata), which are preferentially fed on by Australian brush possum.
southern rata; photo by Stan Shebs
Bernd Blossey, (free access!) who has long studied the role of high deer populations in North American forests, praises the study’s attempt to measure data, not just rely on models, and its inclusion of soil. However, he notes other limitations of the New Zealand study:
The small exclosures (20 x 20 m) are subject to edge effects. Some of Blossey’s exclosures occupy 2 hectares.
Twenty years is too short a time for analysis of such long-term processes as carbon sequestration and regeneration of slow-growing trees. Therefore, any results must be considered preliminary. Furthermore, no one recorded any differences in carbon sequestration of the paired plots at the time the exclosures were set up.
There’s no mention of possible impacts by introduced earthworms.
Dr. Blossey recognizes that the current study’s authors cannot re-do actions taken decades in the past. Still, the data gaps reduce the value of the findings.
I conclude that uncertainties continue due to: the long timelines of species’ regeneration and growth to full sizes; the requirement for large exclosures; the complexity of factors affecting carbon sequestration; and probably other influences.. Managers trying to maximize carbon sequestration are forced to act without truly knowing the best strategy or how their actions will affect the future.
For more about invasive mammals’ impacts in U.S. forests, also see the study by USFS scientists, Poland et al. (full citation listed in sources). One can enter “mammal” in the search box for the on-line PDF.
SOURCES
Allen, K., P.J. Bellingham, S.J. Richardson, R.B. Allen, L.E. Burrows, F.E. Carswell, S.W.Husheer, M.G. St. John, D.A. Peltzer, M. Whenua. 2023. Long-term exclusion of invasive ungulates alters tree recruitment and functional traits but not total forest carbon. Ecological Applications. 2023; e2836. https://onlinelibrary.wiley.com/r/eap
Poland, T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019), Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. The on-line version as at https://link.springer.com/book/10.1007/978-3-030-45367-1
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
This blog asks YOU!!! to support funding for some of the key USDA programs. This blog focuses on USDA’s Animal and Plant Health Inspection Service (APHIS). APHIS is responsible for preventing introduction of pests that harm agriculture, including forests; and for immediate efforts to eradicate or contain those pests that do enter. While most port inspections are carried out by the Department of Homeland Security Bureau of Customs and Border Protection, APHIS sets the policy guidance. APHIS also inspects imports of living plants.
Please help by contacting your members of the House and Senate Appropriations Committees. I provide a list of members – by state – at the end of this blog. APHIS is funded by the House and Senate Appropriations Subcommittees on Agriculture and Related Agencies. These Subcommittees have scheduled hearings on the topic and I’ve drafted written testimony for them. I expect CISP will be joined by additional members of the Sustainable Urban Forest Coalition in signing the testimony. You can add the crucial voice of constituent’s support.
I will blog soon about funding for USDA’s Forest Service (USFS) – I don’t yet have necessary information to suggest specific funding levels.
Your letter or email need be no more than a couple paragraphs. To make the case for greater funding, feel free to pick-and-choose from the information that follows. Your greatest impact comes from speaking specifically about what you know and where you live.
These are the specific dollar amounts we’d like you to ask for. The rationale for each is below.
Appropriations for APHIS programs (in $ millions)
Program
FY 2022 (millions)
FY 2023
FY 2024 Pres.’ request
Our ask
Tree & Wood Pest
$61
$63
$64
$65 M
Specialty Crops
$210
$216
$222
$222 M
Pest Detection
$28
$29
$30
$30 M
Methods Development
$21
$23
$23
$25 M
The Costs of Introduced Pests
Introduced pests threaten many forest products and services benefitting all Americans, including wood products, wildlife habitat, carbon sequestration, clean water and air, storm water management, lower energy costs, improved health, aesthetic enjoyment, and related jobs. Already, the 15 most damaging non-native pests threaten at least 41% of forest biomass in the “lower 48” states. In total, these 15 species have caused an additional annual conversion of live biomass to dead wood at a rate similar in magnitude to that attributed to fire (5.53 TgC per year for pests versus 5.4 to 14.2 TgC per year for fire) [Fei et al.; full citation at end of blog; see also earlier].
tanoaks killed by SOD; Oregon Department of Forestry photo
These pests also impose significant costs that are borne principally by municipal governments and homeowners. As more pests have been accidentally introduced over time, these costs have risen. A study published last year [Hudgins et al.] projected that by 2050 1.4 million street trees in urban areas and communities will be killed by introduced insect pests. Municipalities on the forefront include Milwaukee and Madison Wisconsin; the Chicago area; Cleveland; and Baltimore, Towson, and Salisbury, Maryland. Removing and replacing these trees is projected to cost cities $30 million per year. Additional urban trees – in parks, on homeowners’ properties, and in urban woodlands – are also expected to die and require removal and replacement.
Pathways of Introduction
Tree-killing pests are linked to the international supply chain. Many pests—especially the highly damaging wood-borers like emerald ash borer, Asian longhorned beetle, polyphagous and Kuroshio shot hole borers, and redbay ambrosia beetle—arrive in inadequately treated crates, pallets, and other forms of packaging made of wood. Other pests—especially plant diseases like sudden oak death and sap sucking insects like hemlock woolly adelgid—come on imported plants. Some pests take shelter, or lay their eggs, in or on virtually any exposed hard surface, such as steel, decorative stone, or shipping containers.
infested wood from a crate; Oregon Department of Agriculture photo
Wood Packaging
Imports from Asia have historically transported the most damaging pests, e.g., Asian longhorned beetle, emerald ash borer, redbay ambrosia beetle, and the invasive shot hole borers. For decades goods from Asia have dominated imports. As of February 2022, U.S. imports from Asia were running at a rate of 20 million shipping containers per year. A recent analysis [Haack et al.; see also here] indicates that at least 33,000 of these shipping containers, perhaps twice that number, are carrying a tree-killing pest. These facts have led scientists to project [Leung et al.] that by 2050, the number of non-native wood-boring insects established in the US could triple. Hudgins et al. say the greatest damage would occur if an Asian wood-boring insect that attacks maples or oaks were introduced. Such a pest could kill 6.1 million trees and cost American cities $4.9 billion over 30 years. The risk would be highest if this pest were introduced to the South – and U.S. southern ports are receiving more direct shipments from Asia after the expansion of the Panama Canal in 2016. https://www.nivemnic.us/?m=202207
After introduction of the ALB, APHIS acted to curtail further introductions in wood packaging from China. First – in 1998 – APHIS required China to treat its wood packaging. Second, it worked with foreign governments to develop the International Standard for Phytosanitary Measures (ISPM) #15. The U.S. and Canada began phasing in ISPM#15 in 2005 with full implementation in 2006. Under ISPM#15, all countries shipping goods to North America must treat their wood packaging according to specified protocols with the goal of “significantly reducing” the risk that pests will be present.
However, as I have often blogged [see blogs under “wood packaging” category on this site] ISPM#15 has fallen short. Haack et al. found that as recently as 2020, 0.22% [1/5th of 1 percent] of the shipping containers entering the U.S. were infested by a tree-killing insect. This equates to tens of thousands of containers harboring tree-killing insects.
Worse, the data indicate that our trade partners’ compliance with the rules has deteriorated; the “approach rate” of pest-infested wood packaging fell in 2005-2006, but has since gone back up.
The most troubling offender is China. Although required since 1998 to treat its wood packaging, China consistently has one of the highest pest approach rates: it was 0.73% [or ¾ of 1%] during the 2010-2020 period. This is three times the global average for the period. Since China supplied 40.7% of U.S. imports in 2022 [Szakonyi], or 5,655,000 containers. Thus China alone might be sending to the U.S. 30,000 containers infested with tree-killing insects. These pests threaten our urban, rural, and wildland forests and reduce forest productivity, carbon sequestration, the rural job base, water supplies and quality, and many other ecosystem services.
ISPM#15 falls short at the global level. The fact that a pallet or crate bears the mark indicating that it complies with ISPM#15 has not proved to be reliable.
You might ask your Member of Congress or Senators to ask APHIS what steps it will take to correct the problem of Chinese non-compliance. (Remind him or her that that the Asian longhorned beetle, emerald ash borer, and many other insects of so-far lesser impact were introduced in wood packaging from China.
Asian longhorned beetle
Remind them also that the Department of Homeland Security’s Bureau of Customs and Border Protection has twice enhanced its enforcement of wood packaging rules. In 2017 it began penalizing importers of non-compliant wood packaging under Title 19 United States Code (USC) §1595a(b) or under 19 USC §1592. In 2021, it incorporated the wood packaging requirements into its voluntary C-TPAC program.)
You might also urge them to ask APHIS what steps it is taking at the global level to improve the efficacy of ISPM#15 – or to replace it if necessary to ensure that pests are not being introduced.
spread of beech leaf disease
Imported Plants (“Plants for Planting”)
Some pest types—especially plant diseases like sudden oak death and sap-sucking insects like hemlock woolly adelgid—come on imported plants. The U.S. imported about 5 billion plants in 2021 [MacLachlan]. Recent introductions probably via this pathway include several pathogens — Phytophthoras, rapid ʻōhiʻa death in Hawai`i, beech leaf disease (established from Ohio to Maine), and boxwood blight. Insects have also been introduced on imported plants recently; one example is the elm zigzag sawfly (present in North Carolina, Virginia, and New York and Ontario). https://www.nivemnic.us/?p=4115
An analysis of data from 2009 [Liebhold et al.] found that approximately 12% of plant shipments were infested by a pest. This pest approach rate is more than 50 times higher than the 0.22% approach rate for wood packaging. APHIS has adopted several changes to its phytosanitary system for imported plants in the decade since 2009. A few studies have been published, but they have focussed on insects and excluded pathogens. We have noted that pathogens continue to be introduced via the plant trade. Therefore, please ask your Member or Senators to ask APHIS to facilitate an independent analysis of the efficacy of the agency’s current phytosanitary programs to prevent introductions of pests on important plants, with an emphasis on introductions of plant pathogens.
APHIS is responsible for preventing spread of the SOD pathogen, Phytophthora ramorum, through trade in nursery plants. In recent years California has had few detections in nurseries and little expansion in forests – but the situation suggests that this good news is probably more the result of the drought than of program efficacy. In cooler, wetter conditions in Oregon and Washington, detections in nurseries and alarming detections in the forest or plantings continue.
In 2022, the APHIS SOD Program supported detection and regulatory activities in 25 states. P. ramorum was detected at 18 establishment, 12 of which were first-time detections. The California nursery regulatory program – which is funded by APHIS – saw reduced funding in 2022. We think these cuts are unwise since this year’s very wet winter will probably lead to a new disease outbreaks. Programs in Oregon and Washington continue to detect infestations in additional retailers brought in by plants bought from other nurseries. Washington responded to four separate “trace forward” incidents, one involving more than 160 residential sites. Clearly, the federal-state program is not succeeding in eradicating P. ramorum from nurseries. Please suggest that your Congressperson and Senators ask APHIS what steps it is taking to improve the efficacy of the SOD program.
SOD-infected rhodoendron on plants in Indiana; photo by Indiana Department of Natural Resources
In the East, P. ramorum was found in three of 65 streams sampled in 10 states in 2022 (reaching across the Southeast from Mississippi through North Carolina, plus Texas, Maryland, Pennsylvania, and Illinois). One stream is troubling: a first-time detection in South Carolina, with no obvious nursery source. Since stream sampling began, P. ramorum has been detected from eight streams in four states, Alabama, Mississippi, North Carolina, and now South Carolina. The pathogen has been present in some of these streams for more than 10 years.
Oregon faces particularly high risks. Three of the four known strains of P. ramorum are established in Oregon forests. One of them, the EU1 lineage, is more aggressive than the NA1 clonal lineage already present in forests. In addition, the EU1 strain might facilitate sexual reproduction of the pathogen, thus exacerbating Oregon’s struggle to contain the disease.
As we know, introduced pests do not stay in the cities where they first arrived — they spread! Often that spread is facilitated by our movement of firewood, plants, or outdoor household goods such as patio furniture.
The beech trees so important to wildlife conservation in the Northeast are under attack by two pathogens and at risk to an insect. Most alarming is the spread – in a dozen years! — of beech leaf disease DMF from Ohio to Maine. A leaf-feeding weevil is spreading south in eastern Canada. Please suggest that your Member or Senators to ask APHIS what steps it is taking to prevent the weevil’s introduction to the U.S.
‘Ōhi‘a trees make up 80% of the biomass of forests in both wet and dry areas of the Hawaiian archipelago. It is under attack by two diseases caused by introduced pathogens first detected in 2010. ‘Ōhi‘a forests support more threatened and endangered species than any other forest system in the U.S. They also play a uniquely important role in providing other ecosystem services, including water supplies.
Asking for the Money Pest Problems Deserve
To respond effectively to these pests and to the others that will be introduced in coming years, the key APHIS programs identified above must have adequate funds. The funding levels I request – and hope you will support – are lower than I would wish, but everyone expects the Congress to refuse significant increases in funding (see table at beginning of this blog).
The Tree and Wood Pests account supports eradication and control efforts targeting principally the ALB and spongy (= gypsy) moth. Eradicating the ALB normally receives about two-thirds of the funds. The programs in Massachusetts, New York, Ohio, and South Carolina must continue until eradication succeeds.
Oregon detected the EAB in 2022. Although the state and Portland have been preparing for a decade for this eventuality, there will still be significant impacts. Four percent of Portland’s street trees are ash – more than 9,000 trees. Young ash constitute three percent of young trees in parks. Loss of Oregon’s ash will also have severe ecosystem impacts. In Willamette Valley wetlands, ash constitutes up to 100% of the forest trees. Washington and California are also concerned. Indeed, the Hudgins study identified Seattle and Takoma as likely to lose thousands of ash trees. The numerous ash in riparian forests, windbreaks, and towns of North Dakota are also at risk since the EAB is established in South Dakota, Minnesota, and Manitoba.
APHIS manages damaging pests introduced on imported plants or other items through its Specialty Crops program. The principal example is its efforts to prevent spread of the SOD pathogen through the interstate trade in nursery plants. We noted above that this program is not as successful as it should be. We support the Administration’s request for $222 million; however, you might suggest that your Member or Senator urge APHIS to allot adequate funding under this budget line to management of SOD, rapid ʻōhiʻa death pathogens in Hawai`i, and beech leaf disease and elm zig-zag sawfly in the East.
The Pest Detection program is key to the prompt detection of newly introduced pests that is critical to successful pest eradication or containment. The “Methods Development” program enables APHIS to improve development of essential detection and eradication tools.
The Administration’s request include a $1 million emergency fund. This is far below the level needed to respond when a new pest is discovered. Funding constraints have hampered APHIS’ response to past pest incursions.
Please note that many of the members of the Agriculture Appropriations Subcommittee are from states where non-native pests are probably not top of mind. It is important that everyone that knows about these threats communicate with your Member/Senators!!
Members of House or Senate Subcommittees that Fund APHIS
(Names of Senators are italicized)
STATE
MEMBER
APHIS APPROP
HOUSE
SENATE
AK
Lisa Murkowski
X
AL
Jerry Carl Katie Britt
X
X
X
Calif
Barbara Lee David Valadao Josh Harder Diane Feinstein
X X X
X X X
X
FL
Debbie Wasserman Scultz Scott Franklin
X X
X X
GA
Sanford Bishop
X
X
ID
Mike Simpson
X
IL
Lauren Underwood
X
X
KS
Jerry Moran
X
X
KY
Mitch McConnell
X
X
LA
Julia Letlow Ashley Hinson
X X
X X
MD
Andy Harris Chris Van Hollen
X
X
X
ME
Chellie Pingree Susan Collins
X X
X
X
MI
John Moolenaar Gary Peters
X X
x
X
MN
Betty McCollum
X
X
MS
Cindy Hyde-Smith
X
X
MT
Jon Tester Ryan Zinke
X
X
X
NB
Deb Fischer
X
ND
John Hoeven
X
X
NM
Martin Heinrich
X
X
NV
Mark Amodei
X
OH
Marcy Kaptur
X
X
OR
Jeff Merkley
X
X
X
PA
Guy Reschenthaler
X
X
RI
Jack Reed
X
TX
Michael Cloud Jake Ellzey
X
X X
UT
Chris Stewart
X
VA
Ben Cline
X
X
WA
Dan Newhouse Derek Kilmer
X
X X
WV
Shelly Moore CapitoJoe Manchin
X
X X
WI
Mark Pocan Tammy Baldwin
X X
X
X
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
Haack R.A., J.A. Hardin, B.P. Caton and T.R. Petrice .2022. Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Hudgins, E.J., F.H. Koch, M.J. Ambrose, and B. Leung. 2022. Hotspots of pest-induced US urban tree death, 2020–2050. Journal of Applied Ecology
Leung, B., M.R. Springborn, J.A. Turner, and E.G. Brockerhoff. 2014. Pathway-level risk analysis: the net present value of an invasive species policy in the US. Front Ecol Environ 2014; doi:10.1890/130311
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. Frontiers in Ecology.
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Szakonyi, M. 2023. Sourcing shift from China pulls US import share to more than a decade low.
I have blogged often about the pest risk of wood packaging associated with imports from Asia – especially China – and the shift in that risk arising from import volumes and ports at which they are arriving (increasing volumes entering country at ports along Atlantic and Gulf coasts). [See blogs posted on this site, under the “wood packaging” category (listed below the archives by date).] As noted, U.S. imports from Asia are at all-time highs: in the first three months of 2022, they reached 1.62 million TEU (shipping containers measured as twenty-foot equivalents). This was 31.1% higher than in the same period in pre-pandemic 2019 (Mogelluzzo, B. April 22, 2022).
The most recent information (Szakonyi, M. 2023) confirms that U.S. importers are shifting suppliers to countries other than China, primarily because of lengthy shutdowns in Chinese factories linked to the “0 COVID” policy and some U.S. restrictions and tariffs. Over 2022 (full year), China – including Hong Kong – supplied 40.7% of U.S. imports. This is still a huge proportion, but lower than in 2021, when it was 42.4%. The Journal of Commerce calculates that the number of containers coming from China fell by 435,000. At the current rate of infestation in wood packaging from China calculated by Haack et al. 2022, that might mean about 1,200 fewer containers from China with infested wood packaging entering the U.S.
[Explanation of calculations: I divided 435,000 by 2 to convert 20-ft TEU into 40-ft containers that CBP encounters at the ports; multiplied the result by 0.75 – based on the decade-old Meissner estimate of % of containers that have SWPM; then multiplied the result by .0073 because that is infestation rate for China during 2010-2020 period]
This might be progress. China continues to have a terrible record of non-compliant wood packaging 23 years after U.S. and Canada instituted phytosanitary requirements. According to Haack et al. (2022), packaging from China made up 4.6% of all shipments inspected under the terms of their analysis, but 22% of the 180 consignments with infested wood packaging. Thus the proportion of Chinese consignments with infested wood is five times greater than expected based on their proportion of the dataset. The rate of wood packaging from China that is infested has remained relatively steady = 1.26% during 2003–2004, 0.73% during 2010 – 2020. And the insects present belong to the group that causes the greatest damage: longhorned beetles (Cerambycids). Indeed, 78% of beetles in this family that were detected were from China.
There is some good news: some types of goods likely to be enclosed in crates have decreased notably. The proportion of furniture and other home items imported from China has declined from 71.6% of all U.S. imports in 2010 to 52.6% in 2022. As Haack et al. (2022) found, crates are the type of wood packaging where wood pests are most commonly found. While crates constituted only 7.5% of the wood packaging inspected, they made up 29.4% of the infested packaging – or four times greater than their proportion of the dataset.
The pest risk might not be changing significantly, however, because some of the new suppliers are also in Asia. Vietnam’s share of U.S. imports rose from 8.2% to 8.7%. The types of goods most often imported from Vietnam included electronics, shoes, and apparel. The U.S. has already been invaded by insect-pathogen complexes native to Vietnam, Taiwan, and other parts of southeast Asia – e.g., redbay ambrosia beetle and laurel wilt; invasive shot hole borers and Fusarium disease.
U.S. imports from South Korea, mostly electronics and autoparts, climbed from 3.8% to 4.1%. Imports from India also saw a tiny increase – from 3.8% to 3.9%. These shipments were primarily apparel and iron and steel components. These goods prompt concern because wood packaging associated with heavy materials are often infested by insects (Eyre et al. 2018). The Haack et al. (2022) analysis found two interceptions of wood packaging from Vietnam, one from Korea, and three from India.
Besides, the Journal of Commerce notes that shifts in suppliers cannot go far. These countries’ manufacturing capacity and transportation infrastructure are far below those of China (Szakonyi, M. 2023).
In February 2023, U.S. imports from Asia continued to decline from record levels in 2021 and 2022 to 1.09 million TEU. This level still exceeds by 25% the 869,091 TEU recorded in March 2020, at the beginning of the COVID-19 shutdown (Mongelluzzo, March 17, 2023).
[Reminder: higher shares of imports from Asia are going to ports along the Atlantic and Gulf coasts – spreading the risk. See earlier blogs. In early March the Port of Savannah posted an advertisement to the on-line Journal of Commerce, crowing that by July it will complete straightening the river at the Garden City Terminal (the container terminal). This fix will enable Savannah to raise its annual container processing capacity by 1.5 million TEU, to 7.5 million.]
The most hopeful finding is that imports from Mexico jumped 19.2% in the first 11 months of 2022 compared to the same period in 2021. Importers have their reasons: a desire to buy from producers closer to the U.S. market. These motivations have nothing to do with the risk of forest pest introductions. However, we can rejoice because Mexico has greatly improved the pest-infestation rates of its exports since 2009. The rate fell from 0.29% in 2003-2004 to 0.04% in 2010-2020 (Haack et al. (2022).
larval Asian longhorned beetle; Thomas Denholm, NJ Department of Agriculture; Bugwood
I remain outraged that U.S. agencies have not taken effective steps to deal with the nearly 25-year-long problem of Chinese noncompliance with our phytosanitary requirements. As I noted in my previous blog, link to blog 303 Customs and Border Protection officials are disappointed that their enhanced enforcement in 2017 and 2021 has not yet resulted in improved compliance.
I suggested that the U.S. and Canadian government agencies should penalize trade partners with high records of not complying with ISPM#15. Among steps they should consider are
U.S. and Canada should refuse to accept wood packaging from foreign suppliers that have a record of repeated violations – whatever the apparent cause of the non-compliance. Institute severe penalties to deter foreign suppliers from taking devious steps to escape being associated with their violation record.
APHIS and CBP and their Canadian counterparts should provide guidance to importers on which foreign treatment facilities have a record of poor compliance or suspected fraud – so they can avoid purchasing SWPM from them. I greatly regret that the death of Gary Lovett might put an end to the voluntary industry program he had been developing, described here.
Encourage a rapid switch to materials that don’t transport wood-borers. Plastic is one such material. While no one wants to encourage production of more plastic, the Earth is drowning under discarded plastic. Some firms are recycling plastic waste into pallets.
Haack et al. 2022 fully describes the methodology used, the structure of USDA’s Agriculture Quarantine Inspection Monitoring (AQIM) program, detailed requirements of ISPM#15, the phases of U.S. implementation, etc. Also see the supplemental data sheet in Haack et al. (2022) that compares the methods used in each analysis.
SOURCES
Eyre, D., Macarthur, R., Haack, R.A., Lu, Y. and Krehan, H., 2018. Variation in inspection efficacy by member states of wood packaging material entering the European Union. Journal of Economic Entomology, 111(2), pp.707-715.
Haack RA, Hardin JA, Caton BP and Petrice TR (2022) Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Frontiers in Forests and Global Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Meissner, H., A. Lemay, C. Bertone, K. Schwartzburg, L. Ferguson, L. Newton. 2009. Evaluation of Pathways for Exotic Plant Pest Movement into and within the Greater Caribbean Region. A slightly different version of this report is posted at 45th Annual Meeting of the Caribbean Food Crops Society https://econpapers.repec.org/paper/agscfcs09/256354.htm
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The characteristic zigzag pattern Picture: Kelly Oten, NC State University.
Guest blog by Kelly Oten, NC State University
The elm zigzag sawfly [EZS; Aproceros leucopoda Takeuchi (Hymenoptera: Argidae)] is the newest invasive forest insect detected in the eastern US. The colloquially-used common name, currently going through the ESA common name approval process, is not only catchy, but perfectly describes this defoliator’s unique feeding damage. As EZS feeds on elm leaves, it weaves a zigzag pattern from the margin of the leaf towards the mid-vein.
An Expansive — and Quickly Growing – Range
Native to East Asia, the first confirmation of EZS in North America occurred in August 2020 in Québec, Canada when an iNaturalist user posted a photo showing the characteristic zigzag defoliation. The observer realized it was potentially EZS and emailed local entomologists in the province who visited the site, collected specimens, and obtained species confirmation through the Canadian Food Inspection Agency Entomology Lab (Martel et al. 2021). However, this detection was not actually the beginning.
Three months before the Canadian detection, the same defoliation pattern was observed in Frederick County, Virginia, USA. Observers suspected EZS, but no specimens were recovered and therefore identification could not be confirmed. A year later, the site was revisited and this time, bingo—specimens were present and confirmed as EZS. Subsequent surveys that summer led to detections in eight additional Virginia counties. At the same time, the telltale defoliation was observed in Lehigh County, Pennsylvania, USA, but no specimen could be recovered for confirmation. In 2022, EZS popped up more widely; four additional states confirmed EZS: Pennsylvania, North Carolina, Maryland, and New York.
map created by Kelly Oten, NCSU
Though new to North America, this insect has a history of invasiveness. First detected in Europe in 2003, it now occupies areas from the United Kingdom and France in the west, to Russia and Kazakhstan in the east (Ashikbayev et al. 2018, iNaturalist 2022).
The strange and unusual biology of elm zigzag sawfly
Like all Hymenopterans, EZS goes through four life stages: egg, larva, pupa, adult. Eggs are laid along leaf margins; after hatching, larvae feed on leaf foliage in a zigzag pattern towards the mid-vein. Older larvae consume the leaf more entirely, leaving behind the mid-vein and thicker lateral veins only. Before pupating, larvae spin a cocoon within which they pupate. Cocoons are seasonally dimorphic; summer pupae (which emerge as adults in 4-7 days) are net-like and attached to leaves or twigs. Overwintering pupae are solid-walled and found in leaf litter or soil. Interestingly, overwintering pupae are not just produced from the last generation of the year. Even early in the summer, overwintering pupae may develop alongside summer pupae. Adults are able to begin oviposition immediately; not only do they not need to feed, but they don’t need to find a mate either! EZS is parthenogenetic, meaning they reproduce without mating. In fact, no male EZS has ever been recorded and it’s believed the species is entirely female.
As elm zigzag sawfly larvae (bottom left on leaf) grow, they feed more wholly on elm leaves. Picture: Kelly Oten, NCSU
The entirety of this life cycle can last ~20-36 days when not overwintering. However, the voltinism of this pest is highly variable. Papp et al. (2018) recorded up to seven generations in a year on lab-reared colonies, but in nature in Europe, anywhere between two and six generations has been recorded (Blank et al. 2010, Mol and Vonk 2015). In Virginia, two generations were recorded in 2021 and 1 in 2022. It is unknown what factors play into the number of generations per year, but it’s clear that it’s highly variable. The ability of EZS to multiply rapidly and have multiple generations per year suggest large populations can build in a relatively short time. In fact, this was observed in North Carolina in 2022 and in Europe several times before. Large populations are capable of severe defoliation and may cause long-term impacts on tree health.
This collective life cycle description is based on Blank et al. 2010, Martel et al. 2021, Martynov and Nikulina 2017, and Wu 2006.
Spread
EZS has astonished many with how it seems to be popping up all over the place in such a short amount of time. Since 2020, it has been detected in five US states and at several sites along the St. Laurence River in the Canadian province. The adults are strong fliers, capable of spreading 45-90 km (~27-55 mi) per year (Blank et al. 2014). Given the fact they’re parthenogenetic, relatively small numbers can disperse to begin new populations. Of perhaps greater concern is the potential for long-range dispersal. In heavy infestations in North Carolina, cocoons were found not just attached to leaves and stems, but also non-living objects, suggesting a possible mechanism of long-range dispersal should they become attached to vehicles or other objects transported long distances. In addition, EZS damage ranges from minor to severe defoliation. When populations are low and feeding is minor, it’s less likely to be detected unless intentional surveys are conducted. This cryptic nature might suggest it’s in more places than we are currently aware.
An elm zigzag sawfly cocoon attached to a fence post. Picture: Kelly Oten, NCSU
So, what’s the big deal?
In short, we don’t know yet. Generally speaking, defoliation by insects causes little long-term harm to tree health but severe and/or repeated defoliation can weaken or sometimes kill a host. In Europe, trees severely defoliated by EZS are typically able to re-leaf but may suffer branch dieback and/or reduced growth (Blank et al. 2010, Zandigiacomo et al. 2011). Also of note, EZS is attacking elm, an already-threatened tree due to widespread mortality cause by Dutch elm disease in the 1900s. Defoliation by EZS could further weaken infected trees or, at the very least, present an additional threat for remaining elms (Blank et al. 2010). While it seems aesthetic damage will be the primary concern with EZS, the potential for long-term tree health impactsin the US is uncertain and should be investigated. For now, anyone that finds EZS or its characteristic defoliation pattern are encouraged to report it to their respective state agriculture or forestry agency.
A row of winged elm (Ulmus alata) in NC were severely defoliated by elm zigzag sawfly. Picture: Kelly Oten, NCSU
[See Faith’s earlier blog about the zigzag sawfly here.]
References
Ashikbayev, N. Z., N. S. Mukhamadiyev, G. Z. Mengdibayeva, M. B. Temirzhanov, and N. K. Kuanyshbaev. 2018. Development of forest entomology in Kazakhstan, pp. 42–47. In T. I. Espolov, K. M. Tireuov, E. I. Islamov, S. B. Baizakov, K. T. Abayeva, E. Z. Kentbaev, and B. A. Kentbaeva (eds.), Actual problems of sustainable development in forestry complex, vol. 2. Aitumar Publishing, Almaty, Kazakhstan.
Blank, S. M., H. Hara, J. Mikulás, G. Csóka, C. Ciornei, R. Constantineanu, I. Constantineanu, L. Roller, E. Altenhofer, T. Huflejt, and G. Vétek. 2010. Aprocerosleucopoda (Hymenoptera, Argidae): an East Asian pest of elms (Ulmus spp.) invading Europe. Eur. J. Entomol. 107: 357–367.
Blank, S. M., T. Köhler, T. Pfannenstill, N. Neuenfeldt, B. Zimmer, E. Jansen, A. Taeger, A.D. Liston. 2014. Zig-zagging across Central Europe: recent range extension, dispersal seed and larval hosts of Aprocerosleucopoda (Hymenoptera, Argidae) in Germany. J. Hymenopt. Res. 41: 57-74.
iNaturalist. Available from https://www.inaturalist.org. Accessed August 2022.
Martel, V., O. Morin, S. Monckton, C. Eiseman, C. Béliveau, M. Cusson, and S. Blank. 2021. Elm zigzag sawfly, Aproceros leucopoda (Hymenoptera: Argidae), recorded for the first time in North America through community science. Can. Entomol. 154: E1.
Martynoz, V. V., and T. V. Nikulina. 2017. Population surge of zigzag elm sawfly (Aproceros leucopoda (Takeuchi, 1939): Hymenoptera: Argidae) in the Northern Ciz-Azov Region. Russ. J. Biol. Invasions 8: 25-34.
Mol, A. W. M., and D. H. Vonk. 2015. De iepenzigzagbladwesp Aproceros leucopoda (Hymenoptera: Argidae), een invasieve exoot in Nederland. Entomol. Ber. 75: 50-63.
Papp, V., M. Ladányi, and G. Vétek. 2018. Temperature-dependent development of Aproceros leucopoda (Hymenoptera: Argidae), an invasive pest of elms in Europe. J. Appl. Entomol. 142: 589-597.
Wu, X. Y. 2006. Studies on the biology and control of Aproceros leucopoda. Plant Prot. 32: 98-100.
Zandigiacomo, P., E. Cargnus, and A. Villani. 2011. First record of the invasive sawfly Aproceros leucopoda infesting elms in Italy. Bull. Insectology 64: 145-149.
nearly dead ash in Shenandoah National Park; photo by F.T. Campbell
I participated in the annual USDA Interagency Invasive Species Research Forum in Annapolis in January 2023; as usual, I learned interesting developments. I focus here on updates re: efforts to protect ash and hemlock
Hopeful Developments re: countering EAB to protect ash
There are hopeful results in both the biocontrol and resistance breeding programs. The overall goal is to maintain ash as a viable part of the North American landscape.
Biocontrol
Juli Gould (APHIS) reminded us that the agency began a classical biocontrol program targetting emerald ash borer (EAB) in 2003 – only a year after EAB had been detected and much earlier than is the usual practice. [Thank you, former APHIS PPQ Deputy Administrator Ric Dunkle!] By 2007 scientists had identified, tested, and approved three agents; a fourth was approved in 2015.
Nicole Quinn (University of Florida) stressed that the egg prarasitoid, Oobius — if it is effective — could prevent EAB from damaging trees. However, it is so small that it is very difficult to sample. One small study demonstrated that Oobius will parasitize EAB eggs laid in white fringe trees (Chionanthus virginicus) as well as in ash. This is important because it means this secondary host is not likely to be a reservoir of EAB.
The numbers
According to Ben Slager (APHIS), more than 8 million parasitoids have been released at 950 sites since the program began in 2007. These releases have been in 418 counties in 31 states, DC, and four Canadian provinces. Still, these represent just 28% of infested counties. Parasitoids have been recovered in 21 states and two provinces.
Rafael de Andrade (University of Maryland) specified that these releases included more than 5 million Tetrastichus in 787 sites; ~2.5 million Oobius in 828 sites in 30 states; ~500,000 Spathius agrili – lately only north of the 40th parallel. Releases of Spathiusgalinae began in 2015; so far ~ 470,000 in 395 sites.
Impact
Several presenters addressed questions of whether the agents are establishing, dispersing, and – most important – improving ash survival. Also, can classical biocontrol be integrated with other management techniques, especially use of the pesticide emamectin benzoate.
Dispersal
Several studies have shown that the four biocontrol agents disperse well (with the caveat that Oobius is very difficult to detect so its status is much less certain).
Implementation considerations
De Andrade found that the longer the delay between the date when EAB was detected and release of Oobius, the less likely Oobius will be recovered. Tetrastichus surprised because the higher the numbers released, the fewer were recovered. He could determine no association between recovery of S. agrili and variations in release regime [numbers released; delay in releasing biocontrol agents; or frequency of releases]. He said it is too early to assess Sp. galinae since releases began only in 2015, but he did see expected relationship to propagule pressure – the more wasps released, the higher the number that were recovered. Sp. galinae did surprise in one way: it seemed to perform better at lower latitudes. De Andrade noted he was working data from less than half of release sites. He asked collaborators to submit data!!!!
Initial signs of ash persistence and recovery
Claire Rutledge (Connecticut Agriculture Experiment Station) determined that
More large trees were surviving in plots where the biocontrol agents were released
EAB density was lower at long-invaded sites
Parasitism rates were similar across release age treatments and release/control plots
Gould focused on protecting saplings so they can grow into mature trees which could be sources of seeds to establish future generations. She noted that there are many “aftermath” forests across the northern United States – those dominated by ash saplings.
In Michigan, at a site of green ash, as of 2015 – 2021, EAB populations are still low, parasitism rate by Tetrastichus and S. galinae high. The percentage of saplings that remained healthy was greater than 80%. There were similar findings in white ash in New York: very low EAB larval density; and more than 70% of ash saplings had no fresh galleries. Gould reported that Tetrastrichus impcts could be detected within three years of release.
So, EAB are being killed by the biocontrol agents combined with woodpecker predation; but in their fourth instar, after considerable damage to the trees.
downy woodpecker in Central Park, NYC. photo by Steven Bellovin, Columbia University
Jian Duan reported on two long-term studies in green & white ash in Michigan and New England. His team used the most labor-intensive but best approach to determine EAB larval mortality and the cause – debarking trees – to determine whether the EAB larva were parasitized, were preyed on by woodpeckers, or were killed by undetermined cause, such as tree resistance, disease, or competition. In Michigan, he linked a crash of EAB population in 2010 was caused by Tetrastichus; EAB tried to recover, but crashed again, due to S. galinae. EAB larval densities had been reduced to 10 / m2. Predation by abundant woodpeckers and the native parasitoid Atanycolus was also important.
In New England, EAB has also declined from 20-30 larvae /m2 to ~ 10 m2.
In Michigan, healthy ash with dbh of larger than 5 inches were much more plentiful in sites where parasitoids had been released. Their survival/healthy rate also was much higher in release sites but the difference declined as years passed. In New England there were growing numbers of healthy trees in 2021-22; (almost none in 2017). Duan conceded that he could not prove a direct link but the data points to recovery.
Tim Morris (SUNY-Syracuse) found that white ash saplings continued to die in large numbers, but the mortality rate was significantly below the rate in 2017. Canopy conditions varied; some trees that were declining in 2013 were recovering in 2017. Forty percent of “healthy” ash in 2013 continued recovering in 2021. Few living trees were declining; trees were either healthy or dead. He thinks probably a combination of genetics and presence of parasitoids explains which trees recover. Morris also reported some signs of regeneration.
beaver feeding on ash saplings, Fairfax County, Va; photo by F.T. Campbell
At this point, I noted that in parts of northern Virginia, beavers have killed ash saplings. Morris reported finding the same in some sites in New York. Perhaps others have, also; my comment was greeted by laughter.
Theresa Murphy (APHIS) looked at integration of biocontrol and insecticide treatment in urban and natural sites. A study of black and green ash in Syracuse, NY Naperville, IL, and Boulder, CO found continued high parasitism by Tetrasticus and S. galinae and woodpecker attacks in trees treated with emamectin benzoate. Researchers could not detect Oobius. By 2020, most of the untreated trees had died but treated trees remained healthy.
Murphy has begun studying integration of biocontrol and pesticides in green and black ash forests. The goal is to protect large trees to ensure reproduction; the biocontrol agents do not yet protect the large trees. This is especially important for black ash because it declines very quickly after EAB invades. Sites have been established in New York, through collaboration with New York parks, Department of Environmental Conservation, and the Mohawk tribe. She is still looking for sites in Wisconsin – where EAB is spreading more slowly than expected.
1 of the infested ash in Oregon; photo by Wyatt Williams, ODF
Max Ragozzino of the Oregon Department of Agriculture reported on imminent release of biocontrol agents targetting the recently detected outbreak there. I am encouraged by the rapid response by both the state and APHIS.
EAB resistance in ash
Jennifer Koch (USFS) said the goal is not to produce populations where every seedling is fully EAB-resistant, but to develop populations of ash trees with enough resistance to allow continued improvement through natural selection while retaining sufficient genetic diversity to adapt to future stressors (changing climate, pests, diseases). The program has developed methods to quantify resistance in individuals.. Initial field selections of “lingering ash” were shown to be able to kill as many as 45 % of EAB larvae. Already green ash seedling families have been produced by breeding lingering ash parents. This first generation of progeny had higher levels of resistance, on average, than the parent trees. Each generation of breeding can increase the proportion of resistance. Although the bioassays to test for EAB-resistance are destructive (e.g., cutting and peeling to count numbers of surviving larvae), the potted ash seedling stumps can resprout. Once the new sprouts are big enough they are planted in field trials to correlate bioassay results with field performers. Poor performers are culled; those with higher levels of resistance remain and become sources of improved seed.
To ensure preservation of local adaptive traits, this process must be repeated with new genotypes to develop many seed orchards from across the species’ wide range. To support this work, concerned scientists are building multi-partner collaborative breeding networks. These organizations provide ways for citizens and a variety of partners to engage through monitoring and reporting lingering ash, making land available for test planting, and helping with the work of propagation.
Resistance levels in some of the first generation progeny were high enough for use in horticulture, where it is important that trees can remain healthy in challenging environments (street trees, city parks, landscaping, etc.). Koch hopes to develop about a dozen cultivars comprising the best-performing trees, appropriate for planting in parts of Ohio, Michigan, Indiana, and Pennsylvania. Local NGO partners are planting some of these promising genotypes in Detroit to see how they withstand EAB attack.
a black ash swamp; photo via Flickr
The threat to black ash is especially severe, and this species presents unique difficulties. While scientists found several seedlings from unselected seedlots had killed high levels of larvae, those deaths did not always result in better tree survival. Koch thinks the tree’s defense response becomes detrimental to tree by blocking transport of water and nutrients. She is working with experts in genomics and others, such as Kew Royal Botanic Gardens, to try to identify candidate trees for breeding programs. The genomics work has been supported by APHIS and the UK forest research agency, DEFRA. Michigan and Pennsylvania have supported the breeding work. USFS Forest Health Protection has supported work with black and Oregon ash (see below) (J. Koch, USFS, pers. comm.).
Koch has also begun working with Oregon ash, in collaboration with the USFS Dorena Genetic Resource Center (located in Cottage Grove, Oregon) and other partners.
dead hemlock in Massachusetts; photo by Ian Kinahan, University of Rhode Island
Scientists are still trying to find the right combination of biocontrol, chemical treatments, and silvicultural manipulation.
For several years, hope has focused on two has been on two predatory beetles, Laricobius nigrinus and L. osakiensis. Scott Salom (Virginia Tech) reports that release of these beetles over the past 20 years has had a significant impact on HWA density and tree photosynthetic rate and growth. However, Laricobius aredifficult to rear and they attack only the sistens generation of the adelgid. Ryan Crandall (University of Massachusetts) reports it has been difficult to establish these beetles in the Northeast. He links this difficulty is caused by temporary drops in HWA populations after cold snaps.
Scientists now agree that need to find predators that attack HWA during other parts of its lifecycle. Hope now focuses on silverflies — Leucotaraxis argenticollis and Le. piniperda. While both species are established in eastern North America, the clades in the east feed almost exclusively on pine bark adelgid, and have not begun attacking HWA. Biocontrol practitioners therefore collect flies in the Pacific Northwest for release in the east. Salom is increasing his lab’s capacity to rear silverflies and exploring release strategies.
Preliminary evidence indicates that the western clades of Leucotaraxis are establishing, although data are not yet definitive (Havill, USFS).
Detecting the presence of biocontrol agents presents several challenges. Tonya Bittner (Cornell) described efforts to use eDNA analysis for this. Some puzzles have persisted; e.g., at some sites, she detected eDNA but caught no silverflies. This raised the question of long eDNA associated with the original release might persist. Another problem is that the assay cannot separate the introduced western L. nigrinus from the native congener, L.rubus (which also does not feed on HWA). She continues efforts to improve this technique.
Others explored interactions of the biocontrol agents with insecticides. Salom is studying the impact of soil-applied insecticides on Laricobius populations, which aestivate in the soil. Preliminary results showed significant reduction in the beetle’s population under soil drench application but not under soil injection. He has not yet analyzed all the data.
Michigan is trying to prevent spread of HWA from five counties along the eastern shore of Lake Michigan (where HWA was introduced on nursery stock) to widespread hemlock forests in northern part of the state. Phil Lewis (APHIS) is studying persistence of systemic insecticides in hemlock tissues, particularly twigs and needles. The pesticides involved are imidacloprid, dinotefuran, and Olefin. He has found that pesticide levels are highest 18 – 22 months after treatment, then decline. They are significantly higher after trunk injection compared to bark spray or soil treatments. Imidacloprid had higher residues in twigs; dinotefuran in needles. This difference affects the likelihood of adelgids actually ingesting the toxin.
healthy hemlock in experimental gap; Jefferson National Forest, VA; photo by Bud Mayfield, USFS
Bud Mayfield (USFS) reported on his study of silvicultural strategies to support healthier hemlocks. While hemlocks normally thrive in shade, it has been determined that sunlight assists small trees reducing HWA sufficiently to counter the tree’s leaf-level stress. Small sapling hemlocks grown in sunlight fix more carbon and convert it to growth in shoots and trunk diameter.
Mayfield found promising immediate suppression of HWA in large gaps in Georgia and Tennessee. By the third year the saplings were still growing, although their faster growth had attracted more HWA. These findings were less clear farther north in central Virginia and western Maryland – Mayfield thinks because HWA pressure there is lower. However, managers must maintain the gaps by cutting rapidly-growing competing woody species. He plans to test this strategy farther north in Pennsylvania. He is still trying to determine the optimal size of the gap.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm