Oregon ash in swamp in Ankey National Wildlife Refuge, Willamette Valley, Oregon; photo by Wyatt Williams, Oregon Department of Forestry
1) Funds still not released
As of the end of June, the Office of Management and Budget has not released funds to programs under the USDA Forest Service’ State, Private, and Tribal section. This includes many programs – grants, etc. – that support state and other entities’ efforts and operation of the Forest Health Protection program. Meanwhile, tree-killing insects, pathogens, and nematodes pursue their lives … killing trees in the process.
Congress has not yet acted on legislation that will determine the funding level for USFS FHP and Research programs in Fiscal Year 2026, which begins on October 1. I remind you that the Administration has proposed 0 funds for these programs. Take advantage of the Congressional delay – contact your Member of the House and Senators.
2) the Mediterranean oak beetle (MOB) Xyleborus monographus has now been detected in nine California counties, including Napa, Sonoma, Lake, Sacramento, El Dorado, Yolo, Mendocino, and as of last month, Marin. MOB is also present in Oregon – in Multnomah, Clakamass, Marion and Washington counties.
There is no treatment for infected trees. California authorities urge landowners to search for the insect and remove infected trees – and to avoid moving infested wood.
3) Oregon Department of Forestry has announced that thousand canker disease (TCD) of walnut is killing trees of the Juglans genus in the Willamette Valley. Where the insect has been found in traps, the majority of black walnut trees have since died. Black walnut (Juglans nigra) is not native in Oregon; its range east of the Great Plains. Apparently the range of northern California black walnut (Juglans hindsii) also does not extend into Oregon.
Cities in Oregon are preparing for the inevitable arrival of the emerald ash borer (EAB) (Agrilus planipennis) which was detected in Forest Grove in June 2022. The City of Salem inventoried all of its street & park trees in the last 5 years. It plans to inject a systemic pesticide into at least 550 trees this year and a similar number next year. These trees have been judged to be in good condition. Ash trees in poor condition along streets or in parks are gradually being removed and replaced.
Salem also plans to inventory Oregon ash growing in the city’s natural areas so as to understand where they will need to plant other native species. I blogged earlier about the threat EAB and MOB pose to western Oregon’s wetlands and oak savannahs.
4) Ann Hajek of Cornell and colleagues haves published a review of 20 years of research on entomopathogens that might contribute to control efforts targetting the Asian longhorned beetle (ALB) (Anoplophora glabripennis). The authors call for renewed efforts to find appropriate control agents and techniques. They conclude that various pathogens – especially fungi – can support ALB eradication efforts. They would be particularly helpful if ALB populations spread – or a new outbreak is detected. (Remember, ALB has been detected in seven locations in North America – some the result of more than one introduction; and nine locations in Europe.)
The article is open access!! See
Ann E. Hajek, A.E., E.H. Clifton, and L.F. Solter. 2025. Entomopathogens for control of Asian longhorned beetles (Coleoptera: Cerambycidae). Environmental Entomology, XX(XX), 2025, 1–10 https://doi.org/10.1093/ee/nvaf016
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Northern red oak – one of the species planted in Europe & China as part of sentinel garden project; photo by F.T. Campbell
During the USDA Interagency Forum on Invasive Species, Dr. Eliana Torres Bedoya, from the Bonello lab at Ohio State, provided insights gained from a sentinel garden project operating for the last five years.
The gardens were established in six locations: two in China in the Nanjing area, one each in Italy, Sweden, Ohio, and New Hampshire. The network required collaboration among scientists in several countries, a difficult task in itself. (Jiri Hulcr of the University of Florida has also stressed the importance of mutually beneficial collaborations.)
The focus was on detecting and identifying novel fungal pathogens abroad before they ever enter a country, in an approach called ex patria sentinel plantings. Altogether, 32 tree species were planted in at least one location. For example, Chinese and European tree species were planted in the U.S. to identify potential threats to China and Europe. Conversely, North American species were planted in Europe and China to detect potential threats to the U.S. As noted, the reciprocity is crucial to establishing and maintaining a long-term relationship.
Key information gained to date:
While the scientists isolated several potential pathogens from symptomatic plants, analysis of all plants’ leaf microbiomes showed that asymptomatic plants harbored many more potential pathogens that had not been isolated.
Healthy plants tend to harbor larger and often more diverse microbial communities. This study found that asymptomatic plants supported a significantly more abundant, richer, and taxonomically diverse leaf-associated fungal community than symptomatic plants. Importantly, this pattern pertains also to the subset of taxa classified as potential plant pathogens.
Detection of the full range of fungal pathogens requires that samples must be collected both early and mid-to-late in the growing season because microbes present differ.
Core leaf microbiomes were associated with specific tree species, no matter where they were planted. However, the constituents of the core microbiome were outnumbered by other organisms driven primarily by the location of the planting. This had been expected.
Other contributing factors – in declining order – were geographic location, tree species, season, and host health status. In other words, the phylogenetic relationship.
The drivers of fungal community composition interact in complex ways. For instance, the effect of the plant’s health on pathogenic fungal communities might depend on the host species. This relationship can be further modulated by seasonal variation and geographic context.
European & Asian trees planted in Ohio as part of the sentinel garden program; photo by P (E) Bonello
Implications:
Sentinel gardens can facilitate identification of novel host-pathogen interactions in symptomatic and asymptomatic plants, so they should be adopted / supported by governmental and regional phytosanitary agencies.
The findings demonstrate the need to expand surveillance beyond symptomatic plants – at both sentinel gardens and plant health border inspection stations. Phytosanitary agencies should employ both full microbial community molecular characterization to detect threats in asymptomatic plants and traditional symptom-based approaches. These modern approaches are described in Munck and Bonello 2018 (full reference at end of the blog).
Enrico Bonello (pers. comm.) thinks it is likely that similar context-dependent interactions among host and fungus species, season, and geography also drive disease infection and virulence.
Eliana Torres Bedoya (pers. comm.) clarifies that the leaf microbiome is the community of microorganisms living on and within tree leaves. These microbes can contribute to protecting trees against pathogens, enhance tolerance to environmental stressors such as drought or pollution, and influence how trees interact with their surroundings. Because the composition of the leaf microbiome responds to changes in climate, location, and tree species, it also serves as a valuable indicator of forest health and environmental change.
There are several approaches to studying microbial communities in leaves. One is the traditional, culture-based method, which relies on isolating and cultivating microorganisms on nutrient media. While this approach is effective for recovering fast-growing and easily culturable taxa, it has a major limitation: the vast majority of environmental microbes are not readily culturable under standard laboratory conditions. As a result, full understanding requires use of culture-independent methods. One technique widely used is metabarcoding. This technique involves extracting total DNA from leaf tissue and amplifying a phylogenetically informative genetic marker specific to the microbial group of interest (for example, the internal transcribed spacer (ITS) region for fungi or the 16S rRNA gene for bacteria). The amplified regions are then sequenced using high-throughput sequencing platforms. After a series of processing steps, the sequences are clustered into Amplicon Sequence Variants (ASVs), which represent unique DNA sequences that can be used as proxies for microbial taxa present in the sample. Torres Bedoya and Bonello used the ASVs for comparative analysis.
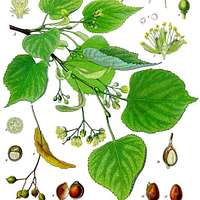
Tilia cordata (linden) via Picaryl (seed wings make a great tea!)
In her presentation, Torres Bedoya provided examples of the complexities arising when trying to detect fungi associated with trees. Eleven potentially pathogenic fungal genera were found to be more abundant in asymptomatic Northern red oak (Quercus rubra) trees (a North American species) planted in both Europe and China. Five ASVs were more abundant in asymptomatic Fraxinus mandshurica trees (an Asian species) planted in Sweden. In this case, the season when the leaves were sampled explained a higher proportion of the variance in the community composition than did the health status of the host. Molecular methods detected 10 genera not revealed through isolation from little-leaf linden species (Tilia cordata) trees (a European species) planted in China and the US.
This “proof of concept” study considered only fungi associated with leaves. As shown above, learning the true plant health risk associated with any tree taxon’s leaves is already complicated and resource-demanding. To fully exploit the power of the ex patria sentinel plantings approach, phytosanitary officials must provide additional resources (land, people, equipment, money) to enable screening of all plant parts, above and below-ground, and all potentially pathogenic taxa, including nematodes, phytoplasmas, and viruses. These systems must be maintained over years.
Reference
Munck, I.A., Bonello, P. 2018. Modern approaches for early detection of forest pathogens are sorely needed in the United States. Forest Pathology 48 (5). doi:10.1111/efp.1445
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
ash tree killed by emerald ash borer; photo courtesy of (then) Mayor of Ann Arbor John Hieftje
Since the 1990s, scientists have been trying to the determine costs imposed by invasive species. They hope that measuring monetary costs will motivate political decision-makers to take more assertive actions to counter this ecological treat. As Daigne et al. (2021) point out, too few countries are implementing effective control and mitigation strategies. They say this inaction stems, largely, from undervaluing bioinvasions’ impacts by the general public, stakeholders and decision-makers.
A major step in this effort was creation of the InvaCost database. The goal was to provide a reliable, comprehensive, standardized and easily updatable synthesis of bioinvasions’ monetary costs worldwide.
Several publications based on this database appeared. I have blogged about studies published in 2021 or 2022: a) the costs of bioinvasions generally (Cuthbert et al. 2022); b) the costs imposed by invasive species in protected areas (Moodleyet al. 2022; c) a focus on the “worst” 100 invasives (as determined by the IUCN) (Ahmedet al. 2022); and d) assessing costs associated with various pathways of introduction (Turbelin et al. 2022).
The InvaCost database, as applied in these studies, demonstrated that bioinvasions impose tremendous costs –a minimum of US $1.288 trillion for the period 1970 – 2017. These costs increased on average three times per decade (Daigne et al. 2022).
Still, everyone has recognized that InvaCost data have significant limitations. First, three-quarters of the records in the original database came from North America, Oceania and Europe; and referred to animal taxa, even though plants are a major group of invaders. Also, a large proportion of total invasion costs – for all taxa – probably is undetected. Finally, the many non-market values of species and ecosystems are extremely difficult to calculate (Daigne et al. 2022).
As a result of these deficiencies, the earlier studies discussed in the blogs referenced above substantially underestimated the true costs associated with bioinvasion (Cuthbert et al. 2022).
Now a new study, led by Ismael Soto, finds that the underestimate is huge. Global costs associated with a subset of 162 species (17% of all the species in the InvaCost database) is nearly 17 times higher than reported in the InvaCost database.
Soto et al. (2025) applied species distribution models and macroeconomic data to interpolate these 162 species’ probable impacts in 172 countries
Japanese knotweed – one of the invasive plants proving very costly in Europe, according to I. Soto
The newly identified costs were greatest in Europe; second place fell to North America. This is because both higher damage costs and management expenditures are linked to higher gross domestic product and extent of agricultural area, in addition to environmental suitability. Analysis of monetary costs per unit area revealed that ‘cost hot spots’ are predominantly located in densely populated urban areas and locations hosting key industries. These tend to be in coastal zones, i.e., Europe, the east coast of China, and the east and west coasts of the US.
cypress aphid Cinara cupressi – a threat to both native & plantation trees in Africa; photo by Blackman & Eastop via Wikimedia
The authors found that the greatest increase in estimated costs for countries in Africa and Asia. These countries had not previously recorded any economic costs arising from invasions by these 162 species. I have blogged about forest pest threats in Africa.
The authors also significantly increased estimated costs linked to invasive plants. Daigne et al. found that invasive insects caused ~90% of reported costs in the InvaCost database as of 2022. Vertebrates ranked second, plants third. In contrast, Soto et al. determined that invasive plants had the highest average estimated damage costs (US $42.10 billion) and management expenditures ($0.81 billion).
Substantial total costs were also reported for arthropods, mammals and birds. Reported damage and management costs were much lower for molluscs, fish, reptiles and amphibians. Daigne et al. suggest this might be due to their lower (observable) damage to human infrastructure, research biases leading to fewer studies, or disparities resulting from the filtering process used in their own study.
Williams et al. (2023) focus on insects, which cause damage primarily to agriculture, human health, and forestry. Insects constitute the highest number of species introduced as ‘Contaminants’ (n = 74) and ‘Stowaways’ (n = 43). They also impose the highest costs among species using these two pathways.
Forest insects and pathogens account for less than 1% of the records in the InvaCost database. I believe that this figure reflects significant under-reporting of these invasion events. Even at this paltry level of reported invasions, forest insects and pathogens were responsible for causing 25% of total annual costs ($43.4 billion) (Williams et al. 2023). This discrepancy illustrates the huge economic cost associated with widespread mortality of trees. Yet authorities in most countries continue to provide completely inadequate resources to counter this threat.
The authors of these publications examining economic losses associated with bioinvasion all note that ecological damage is additional. Soto et al. note that bioinvasions contribute to 60% of already recorded global extinctions. Interestingly, the species ranked third using the criterion of monetary damage is the cactus moth Cactoblastis cactorum. This insect threatens flat-padded Opuntia cacti across the United States and in the center of endemism, Mexico.
a flat-padded Opuntia — vulnerable to the cactus moth; photo by F.T. Campbell
Soto et al. found a lag of ~46 years between first (reported) detection of an introduced species and the peak of damage costs. They suggest that the rising monetary cost reflects the species becoming more abundant or occupying a larger area. The authors also say this finding demonstrates the value of implementing mitigation measures as soon as possible. Their finding thus validates others’ advocacy for investing in prevention and rapid response measures (see Cuthbert et al. and Daigne et al.). Soto et al. were cheered by the fact that spending on management measures – when it was reported – often followed soon after a species’ detection – or even before (e.g., prevention).
But Decision-Makers Usually Delay – Why?
Prevention is a hard sell. Decision-makers find it difficult to justify management expenditures before impacts become obvious. By that time, of course, management of the invasion is extremely difficult and expensive – if it is possible at all. Ahmed et al. found particularly effective wording to describe this problem: bioinvasion costs can be deceitfully slow to accrue, so policy makers don’t appreciate the urgency of taking action. Another contributing factor is that when efficient proactive management succeeds in preventing any impact, it paradoxically undermines evidence of the value of this action!
Programs to minimize the economic and ecological consequences of bioinvasion are severely obstructed – if not doomed! – by the following difficulties:
Resources are in short supply. Experts find that demands to address other threats to agriculture or natural systems outcompete appeals to ramp up invasive species efforts.
Prediction is uncertain. Cuthbert et al. found that none of the species with the highest pre-invasion investment was among the top 10 costliest invaders in terms of damages. Cuthbert et al. do not discuss whether this is evidence that the prevention efforts were effective? Or, alternatively, that prevention efforts target the wrong species.
Harm is in the eye of the beholder. Stakeholders’ perceptions of whether an introduced species causes a detrimental impact vary. For example, Moodley et al. point out that species imposing the highest economic costs might not be the ones causing the greatest ecological harm.
Externalities. Those harmed by a bioinvasion often are different from those that decide whether to act. Ahmed et al. argue that this creates a moral dilemma.
These decisions are political — influenced by citizens’ expressed wishes. Changing decision-makers’ perceptions of what is important is up to us!!! Start a parade!!!
SOURCES
Ahmed, D.A., E.J. Hudgins, R.N. Cuthbert, .M. Kourantidou, C. Diagne, P.J. Haubrock, B. Leung, C. Liu, B. Leroy, S. Petrovskii, A. Beidas, F. Courchamp. 2022. Managing biological invasions: the cost of inaction. Biol Invasions (2022) 24:1927–1946 https://doi.org/10.1007/s10530-022-02755-0
Diagne, C., B Leroy, A-C. Vaissière, R.E. Gozlan, D. Roiz, I. Jaric, J-M. Salles, C.A. Bradshaw, and F. Courchamp. 2021. High and rising econ costs of bioinvasions worldwide Published online: 31 March 2021
Moodley, D., E. Angulo, R.N. Cuthbert, B. Leung, A. Turbelin, A. Novoa, M. Kourantidou, G. Heringer, P.J. Haubrock, D. Renault, M. Robuchon, J. Fantle-Lepczyk, F. Courchamp, C. Diagne. 2022. Surprisingly high economic costs of bioinvasions in protected areas. Biol Invasions. https://doi.org/10.1007/s10530-022-02732-7
Soto, I., P. Courtois, A. Pili, E. Tordoni, E. Manfrini, E. Angulo, C. Bellard, E. Briski, M. Buric, R.N. Cuthbert, A. Kouba, M. Kourantidou, R.L. Macêdo, B. Leroy, P.J. Haubrock, F. Courchamp and B. Leung. 2025. Using species ranges and macroeconomic data to fill gap in costs of biological invasions. Nat Ecol Evol doi: 10.1038/s41559-025-02697-5
Turbelin, A.J., C. Diagne, E.J. Hudgins, D. Moodley, M. Kourantidou, A. Novoa, P.J. Haubrock, C. Bernery, R.E. Gozlan, R.A. Francis, F. Courchamp. 2022. Introduction pathways of economically costly invasive alien spp. Biol Invasions (2022) 24:2061–2079 https://doi.org/10.1007/s10530-022-02796-5
Williams, G.M., M.D. Ginzel, Z. Ma, D.C. Adams, F.T. Campbell, G.M. Lovett, M. Belén Pildain, K.F. Raffa, K.J.K. Gandhi, A. Santini, R.A. Sniezko, M.J. Wingfield, and P. Bonello. 2023. The Global Forest Health Crisis: A Public Good Social Dilemma in Need of International Collective Action. Annual Review of Phytopathology Vol. 61, 2023
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
In early May I posted a blog about the Trump Administration’s proposed budget – saying that it would eliminate funding for nearly all USFS research & Forest Health Protection.
I can now provide some additional information.
The Administration has released a supplemental document providing a few details about the severe cuts it is proposing for USFS programs vital to countering bioinvasion in the coming fiscal year (FY2026), which starts October 1st. You can download this document at https://www.whitehouse.gov/wp-content/uploads/2025/05/appendix_fy2026.pdf
Congress has the final say on appropriations – so please!!! inform your representative & senators about why these cuts are disastrous.
USFS [See pages 162-168 of the Appendix]
Research & Development
The Administration requests $0 for R&D. It says it will strategically use existing carryover balances to responsibly terminate research programs & close research stations. Thus, funding for R&D will decrease from the $301 million in FY24 to $44 million in FY26. The Forest Inventory & Analysis will be shifted to the National Forest System and funded at $21.5 million – less than program supporters are seeking.
The proposal does contain an “additional amount” of $26 million for dealing with the consequences of wildfires, hurricanes & other natural disasters that occurred in calendar years 2022, 2023, and 2024. I am confused about this funding.
State, Private, and Tribal Forests
The Administration requests $0 for S,P&T. Again, the proposal says the agency will use existing carryover balances to effectively & responsibly terminate these programs. The number of employees would be cut from 520 employees in FY24 to 37.
Again, the proposal contains an “additional amount” of $208 million for Forest Health Management to deal with the consequences of wildfires, hurricanes & other natural disasters that occurred in calendar years 2022, 2023, and 2024. $14 million of this sum is earmarked for assistance to states in the Northeast that are anticipating an outbreak of eastern spruce budworm (which has been spreading from Canada). In a highly unusual move, the proposal says this funding is not subject to a requirement that grant recipients provide matching funds from non-federal sources. [Is it a coincidence that Maine Senator Susan Collins chairs the Senate Appropriations Committee?]
National Forest System
Total funding for NFS would be $1.5 billion. This includes an “additional amount” of nearly $2.5 billion for expenses related to the consequences of wildfires, hurricanes & other natural disasters that occurred in calendar years 2022, 2023, and 2024. $75 million of this amount is earmarked for construction or maintenance of shaded fuel breaks in the Pacific Northwest.
As I noted above, the Forest Inventory and Analysis program would be placed under the NFS.
I am particularly concerned that the budget proposal provides explicitly for $20 million to improve or maintain landscape & watershed conditions by preventing invasive plant infestations and installing aquatic organism passages, etc. There is no mention of programs intended to address damage caused by non-native insects and pathogens. It appears that the Administration proposes to drop all programs re: these organisms.
The overall objective of NFS programs is defined as managing the forests for productive use & resilience to catastrophic wildfire & provide broad range of ecosystem services. The budget allegedly prioritizes funding of programs designed to increase health & resilience of National Forests & Grasslands – including meeting multiple use requirements for resources on these lands.
The prose no longer says that timber production is the sole purpose of Nation forests – as the original budget stated.
APHIS appears to have survived – although the supplement provides minimal information (on pp. 85 – 87 of the Appendix).
The supplement contains a lengthy description of APHIS’ purpose — to protect America’s agricultural and natural resources from introduced pests. It requests $1.1 billion for FY2026. The only plant pest listed as a priority is exotic fruit flies. Personnel would be cut from 6,142 in FY24 to 5,092. I could find no specifics regarding funding for programs of interest – tree & wood pests, specialty crops, pest detection, and methods development.
Implications for Non-native Insects and Pathogens
Remember that USFS’s research and development program is intended to improve forest managers’ understanding of ecosystems, including human interactions and influences, thereby enabling improvements to the health and use of our Nation’s forests and grasslands. Most importantly to me, this program provides foundational knowledge needed to develop effective programs to prevent, suppress, mitigate, and eradicate the approximately 500 non-native insects and pathogens that are killing America’s trees.
The Forest Health Program provides technical and financial assistance to the states and other forest-management partners to carry out projects (designed based on the above research) intended to prevent, suppress, mitigate, and eradicate those non-native insects and pathogens. The program’s work on non-federal lands is crucial because introduced pests usually start their incursions near cities that receive imports (often transported in crates, pallets, or imported plants).
[FIA might inform all about where such pests are found — but it doesn’t address how to contain their spread, suppress their impacts, or restore the affected tree species.]
Eliminating either or both programs will allow these pests to cause even more damage to forest resources – including timber.
Both supporting research and on-the-ground management must address pest threats across all U.S. forests, including the more than 69% that are located on lands managed by others than the USFS. Already, the 15 most damaging of these pests threaten destruction of 41% of forest biomass in the “lower 48” states. This is a rate similar in magnitude to that attributed to fire (Fei et al. 2019). [This estimate does not include loss of beech beech leaf disease.] It is ironic that the Administration considers the fire threat to be so severe that it has proposed restructuring the government’s fire management structure.
I remind you that the existing USFS R&D budget allocates less than 1% of the total appropriation to studying a few of the dozens of highly damaging non-native pests. I have argued that this program should be expanded, not eliminated. Adequate funding might allow the USFS to design successful pest-management programs for additional pests (as suggested by Coleman et al.).
As a new international report (FAO 2025) notes, genetic resources underpin forests’ resilience, adaptability, and productivity. Funding shortfalls already undercut efforts to breed trees able to thrive despite introduced pests and climate change (the latter threat is still real, although the Administration disregards it). I have frequently urged the Congress to increase funding for USFS programs – which are sponsored primarily by the National Forest System and State, Private, and Tribal, although some are under the R&D program.
I repeat: Please ask your Member of Congress and Senators to oppose these proposed cuts. Ask them to support continued funding for both USFS R&D and its State, Private, and Tribal Programs targetting non-native insects and pathogens. America’s forests provide resources to all Americans – well beyond only timber production and they deserve protection.
Contacting your Representative and Senators is particularly important if they serve on the Appropriations committees.
House Appropriations Committee members:
Republicans: AL: Robert Aderholt, Dale Strong; AR: Steve Womack; AZ: Juan Ciscomani; CA: Ken Calvert, David Valadao, Norma Torres; FL: Mario Diaz-Balart, John Rutherford, Scott Franklin; GA: Andrew Clyde; ID: Michael Simpson; IA: Ashley Hinson; KY: Harold Rogers; LA: Julia Letlow; MD: Andy Harris; MI: John Moolenaar; MO: Mark Alford; MS: Michael Guest; MT: Ryan Zinke; NC: Chuck Edwards; NV: Mark Amodei; NY: Nick LaLota; OH: David Joyce; OK: Tom Cole, Stephanie Bice; PA: Guy Reschenthaler TX: John Carter, Chuck Fleishmann, Tony Gonzales, Michael Cloud, Jake Ellzey; UT: Celeste Maloy; VA: Ben Cline; WA: Dan Newhouse; WV: Riley Moore
Democrats: CA: Pete Aguilar, Josh Harder, Mike Levin; CT: Rosa DeLauro; FL: Debbie Wasserman Schultz, Lois Frankel; GA: Sanford Bishop; HI: Ed Case IL: Mike Quigley, Lauren Underwood; IN: Frank Mrvan; MD: Steny Hoyer, Glenn Ivey; ME: Chellie Pingree; MN: Betty McCollum; NJ: Bonnie Watson Coleman NY: Grace Meng, Adriano Espaillat, Joseph Morelle; NV: Susie Lee; OH: Marcy Kaptur; PA: Madeleine Dean; SC: James Clyburn; TX: Henry Cuellar, Veronica Escobar; WA: Marie Gluesenkamp Perez; WI: Mark Pocan
Senate Appropriations Committee members:
Republicans: AK: Lisa Murkowski; AL: Katie Britt; AR: John Boozman (AR); KS: Jerry Moran; KY: Mitch McConnell; LA: John Kennedy; ME: Susan Collins; MS: Cindy Hyde-Smith; ND: John Hoeven; NE: Deb Fischer; OK: Markwayne Mullin; SC: Lindsey Graham; SD: Mike Rounds TN: Bill Hagerty; WV: Shelley Moore Capito;
Democrats: CT: Chris Murphy; DE: Chris Coons; GA: Jon Ossof; HI: Brian Schatz; IL: Richard Durbin; MD: Chris van Hollen; MI: Gary Peters; NH: Jeanne Shaheen; NM: Martin Heinrich; NY: Kirsten Gillibrand; OR: Jeff Merkley; RI: Jack Reed; WA: Patty Murray; WI: Tammy Baldwin
Addendum
Maintaining the USFS State, Private, and Tribal (SPT) programs is essential to
complying with laws adopted by the Congress (see second page).
meeting the USFS mission of sustaining“the health, diversity, and productivity of the nation’s forests and grasslands to meet the needs of present and future generations.”
ensuring future economic and ecological benefits to Americans.
More than two-thirds of U.S. forests are privately owned or managed by state, local, or tribal governments. These forests provide many benefits, including 89% of America’s timber harvest.[i] SPT is the only federal program providing technical, financial, & educational assistance to these non-federal landowners.
Among the many threats to American forests, the Center for Invasive Species Prevention (CISP) focuses on the threat from insects and pathogens introduced from abroad. More than 41% of forest biomass in the “lower 48” states is at risk to non-native pests already established in the country.[ii] From 2011 to 2020, sap feeders, e.g., hemlock woolly adelgid, killed trees on 635,000 acres; foliage feeders, e.g., spongy moth, killed trees on 948,884 acres.[iii] Additional pests will be introduced and kill more trees.
Non-native pests are introduced primarily in crates, pallets or other packaging made of wood; and in imported plants. These imports – and the pests – usually land in cities or suburbs and establish there. Initially they cause widespread death of urban trees and impose high costs on local governments and property owners who must remove dying trees. The pests also spread. Hemlock woolly adelgid, emerald ash borer, polyphagous and Kuroshio shot hole borers, goldspotted oak borer, sudden oak death, and beech leaf disease have all spread to National forests from cities or suburbs.
The most effective way to protect America’s forests is to find and kill the pests where they first appear – usually in city trees. Waiting to act until a pest reaches National Forest boundaries means failure. Instead, we should expand the Forest Health Management (FHM) Cooperative Lands program to quickly detect, contain, and – if possible – eradicate the pests. With higher appropriations, the STP FHM program could tackle more of the 53 tree species under threat. At present, only four of these species benefit from 95% of FHM projects – eastern oaks, loblolly and ponderosa pines, and hemlocks.[iv]
USFS Research and Development (R&D) program
FHM adopts strategies based on knowledge of pests’ life histories and traits gained through research conducted or sponsored by the USFS R&D program. CISP urges you to support continued funding for the USFS Research and Development (R&D) program. However, we advocate a realignment: raise the proportion of research funding allocated to invasive species from the current paltry level of 1% to 5%. Funding for studying non-native pests has decreased 70% since FY2010 despite new pests attacking our forests. As a result, the Forest Service is hampered from developing effective programs to prevent, suppress, and eradicate most non-native pests.
Another crucial strategy for reducing loss of tree species to non-native pests is breeding trees able to thrive despite introduced pests. Currently these projects are supported – inadequately – by all three USFS divisions: R&D, SPT, and National Forest System (NFS).
The model program is the Dorena Genetic Resource Center. The Center has bred Western white pine and Port-Orford-cedar trees resistant to introduced pathogens; these trees are now being planted. Promising projects target the pathogens killing whitebark pine, American chestnut, American elm, and Hawaiian koa. Projects at earlier stages address ash, beech, and ʻōhiʻa.
Lesson: federal dollars, wisely invested, can mitigate the damage caused by invasive species. CISP asks you to support continuing these programs so that America can restore threatened trees to our forests.
Complying with the Law
The Cooperative Forestry Assistance Act of 1974
Section 2 (a) Findings …—
(1) most of the productive forest land of the United States is in private, State, and local governmental ownership, and the capacity of the United States to produce renewable forest resources is significantly dependent on such non-Federal forest lands;
(b) Purpose.—… authorize[s] the Secretary …, with respect to non-Federal forest lands … to assist in—
…
(3) the prevention and control of insects and diseases affecting trees and forests;
(c) Priorities.—In allocating funds … , the Secretary shall focus on the following national private forest conservation priorities, …:
…
(2) Protecting forests from threats, including … invasive species, insect or disease outbreak, … and restoring appropriate forest types in response to such threats.
(e) Policy. … it is in the national interest for the Secretary to work through and in cooperation with State foresters, or equivalent State officials, nongovernmental organizations, and the private sector …
Healthy Forests Restoration Act of 2003
Sec. 401(a) FINDINGS.—(1) high levels of tree mortality resulting from insect infestation (including the interaction between insects and diseases) may result in — (A) increased fire risk; … (E) degraded watershed conditions; (F) increased potential for damage from other agents of disturbance, including exotic, invasive species; and (G) decreased timber values;
…
(3) the hemlock woolly adelgid is— (A) destroying streamside forests throughout the midAtlantic and Appalachian regions; (B) threatening water quality and sensitive aquatic species; and (C) posing a potential threat to valuable commercial timber land in northern New England;
(4)(A) the emerald ash borer … has quickly become a major threat to hardwood forests …; and (B) … threatens to destroy more than 692,000,000 ash trees in forests in Michigan and Ohio alone, and between 5 and 10 percent of urban street trees in the Upper Midwest;
…
(11)(A) often, there are significant interactions between insects and diseases; (B) many diseases (such as white pine blister rust, beech bark disease, and many other diseases) can weaken trees and forest stands and predispose trees and forest stands to insect attack; and (C) certain diseases are spread using insects as vectors (including Dutch elm disease and pine pitch canker); …
(b) … The purposes of this title are— (1) to require the Secretary to develop an accelerated basic and applied assessment program to combat infestations by forest-damaging insects and associated diseases; (2) to enlist the assistance of colleges and universities …, State agencies, and private landowners to carry out the program; and (3) to carry out applied silvicultural assessments.
Sec. 402 Definitions
…
(3) FOREST-DAMAGING INSECT. … means … (D) a gypsy moth; (E) a hemlock woolly adelgid; (F) an emerald ash borer; … and (I) such other insects … identified by the Secretary.
[i] Oswalt, S.N., .W.B. Smith, P.D. Miles, & S.A. Pugh. Forest Resources of the United States, 2017 Uport WO-97SDA Forest Service Gen. Tech. Report WO-97. March 2019
[ii] Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
[iii] Coleman, T.W, A.D. Graves, B.W. Oblinger, R.W. Flowers, J.J. Jacobs, B.D. Moltzan, S.S. Stephens, R.J. Rabaglia. 2023. Evaluating a decade (2011–2020) of integrated forest pest management in the United States. Journal of Integrated Pest Management, (2023) 14(1): 23; 1–17
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
The Trump Administration’s budget for Fiscal Year 2026 [which begins at the end of September 2025] proposes to eliminate funding for nearly all USFS research & Forest Health Protection.
Proposed Cuts to USFS Research: Timber the Sole Aim
In a letter from Office of Management and Budget (OMB) to Senate Appropriations Committee Chair Susan Collins (R-Maine, Director Russell Vought says the Administration wants to manage National forests “for their intended purpose of producing timber” and that the research and development program “is out of step with the practical needs of forest management for timber production.” The Administration proposes to eliminate funding for USFS research projects other than the small portion covering Forest Inventory and Analysis.
I understand that the USFS Chief told various NGOs that his job is to run the National Forest System, increase timber production by 40%, and do nothing else.
This single aim conflicts with the 1897 legislation founding and authorizing the USFS. It also violates provisions of subsequent legislation such as the Multiple-Use Sustained-Yield Act of 1960 and the National Forest Management Act of 1976. It also departs from long-standing US Forest Service policy – which is the intention.
The “intended purpose” of establishing “forest reserves” [which were later renamed National forests] has never been solely for timber production. The “Organic Act” of 1897 provided that any new forest reserves would have to meet the criteria of forest protection, watershed protection, and timber production.
Specifically, theORGANIC ACT OF 1897 [PUBLIC–No.2.] says:
“[All public lands heretofore designated and reserved by the President of the US under the provisions of the Act [of] March 3rd 1891, the orders for which shall be and remains in full force and effect, unsuspended and unrevoked, and all public lands that may hereafter be set aside as public forest reserves under said act, [these were the “forest reserves,”predecessors of “National Forests]” shall be as far as practicable controlled and administered in accordance with the following provisions:
“No public forest reservation shall be established, except to improve and protect the forest within the reservation, or for the purpose of securing favorable conditions of water flows, and to furnish a continuous supply of timber for the use and necessities of [US] citizens; but it is not the purpose or intent of these provisions, or of the Act providing for such reservations, to authorize the inclusion therein of lands more valuable for the mineral therein, or for agricultural purposes, than for forest purposes.”
The Department of the Interior, which then managed these forest reserves, promptly issued implementing regulations. The regulations stated that the “object” of forest reservations was:
“2. Public forest reservations are established to protect and improve the forests for the purpose of securing a permanent supply of timber for the people and insuring conditions favorable to continuous water flow.”
Therefore, I think the Administration has exaggerated the emphasis on timber production by calling it “the” intended purpose of the original establishment of National forests. The Administration has also chosen to ignore subsequent legislation, such as the Multiple-Use Sustained-Yield Act of 1960 and the National Forest Management Act of 1976.
Sec. 13 of the NFMA limits the sale of timber from each national forest to a quantity equal to or less than a quantity which can be removed from such forest annually in perpetuity on a sustained-yield basis. This limit might be exceeded under certain circumstances, but such excess must still be consistent with the multiple-use management objectives of the land management plan. Further, Sec. 14 requires public input into any decision to raise timber allowances.
During his period as Chief (1905 – 1910), Gifford Pinchot invented and applied the concept of “conservation” of natural resources. As a result “wise use” became accepted as the national goal.
Culminating more than a century of legislation and informed policy, the mission of the USDA Forest Service is to “sustain the health, diversity, and productivity of the nation’s forests and grasslands to meet the needs of present and future generations.”
Proposed Cuts to State, Private, and Tribal Forests
The budget also cuts $303 million from the State, Private, and Tribal Forests program. (I understand this zeroes out the entire program). The OMB Director alleges that the program has been “plagued by oversight issues, including allegation of impropriety by both the Agency and State governments.” I understand that this would eliminate the cooperative projects managed by the Forest Health Protection program, too.
Implications for Non-native Insects and Pathogens
Remember that USFS’s research and development program is intended to improve forest managers’ understanding of ecosystems, including human interactions and influences, thereby enabling improvements to the health and use of our Nation’s forests and grasslands. Most importantly to me, this program provides foundational knowledge needed to develop effective programs to prevent, suppress, mitigate, and eradicate the approximately 500 non-native insects and pathogens that are killing America’s trees.
The Forest Health Program provides technical and financial assistance to the states and other forest-management partners to carry out projects (designed based on the above research) intended to prevent, suppress, mitigate, and eradicate those non-native insects and pathogens. The program’s work on non-federal lands is crucial because introduced pests usually start their incursions near cities that receive imports (often transported in crates, pallets, or imported plants).
Eliminating either or both programs will allow these pests to cause even more damage to forest resources – including timber.
Both supporting research and on-the-ground management must address pest threats across all U.S. forests, including the more than 69% that are located on lands managed by others than the USFS. Already, the 15 most damaging of these pests threaten destruction of 41% of forest biomass in the “lower 48” states. This is a rate similar in magnitude to that attributed to fire (Fei et al. 2019). It is ironic that the Administration considers the fire threat to be so severe that it has proposed restructuring the government’s fire management structure.
I remind you that the existing USFS R&D budget allocates less than 1% of the total appropriation to studying a few of the dozens of highly damaging non-native pests. I have argued that this program should be expanded, not eliminated. Adequate funding might allow the USFS to design successful pest-management programs for additional pests (as suggested by Coleman et al.).
As a new international report (FAO 2025) notes, genetic resources underpin forests’ resilience, adaptability, and productivity. Funding shortfalls already undercut efforts to breed trees able to thrive despite introduced pests and climate change (the latter threat is still real, although the Administration disregards it). I have frequently urged the Congress to increase funding for USFS programs – which are sponsored primarily by the National Forest System and State, Private, and Tribal, although some are under the R&D program.
Please ask your Member of Congress and Senators to oppose these proposed cuts. Ask them to support continued funding for both USFS R&D and its State, Private, and Tribal Programs targetting non-native insects and pathogens. America’s forests provide resources to all Americans – well beyond only timber production and they deserve protection.
Contacting your Representative and Senators is particularly important if they serve on the Appropriations committees.
House Appropriations Committee members:
Republicans: AL: Robert Aderholt, Dale Strong; AR: Steve Womack; AZ: Juan Ciscomani; CA: Ken Calvert, David Valadao, Norma Torres; FL: Mario Diaz-Balart, John Rutherford, Scott Franklin; GA: Andrew Clyde; ID: Michael Simpson; IA: Ashley Hinson; KY: Harold Rogers; LA: Julia Letlow; MD: Andy Harris; MI: John Moolenaar; MO: Mark Alford; MS: Michael Guest; MT: Ryan Zinke; NC: Chuck Edwards; NV: Mark Amodei; NY: Nick LaLota; OH: David Joyce; OK: Tom Cole, Stephanie Bice; PA: Guy Reschenthaler TX: John Carter, Chuck Fleishmann, Tony Gonzales, Michael Cloud, Jake Ellzey; UT: Celeste Maloy; VA: Ben Cline; WA: Dan Newhouse; WV: Riley Moore
Democrats: CA: Pete Aguilar, Josh Harder, Mike Levin; CT: Rosa DeLauro; FL: Debbie Wasserman Schultz, Lois Frankel; GA: Sanford Bishop; HI: Ed Case IL: Mike Quigley, Lauren Underwood; IN: Frank Mrvan; MD: Steny Hoyer, Glenn Ivey; ME: Chellie Pingree; MN: Betty McCollum; NJ: Bonnie Watson Coleman NY: Grace Meng, Adriano Espaillat, Joseph Morelle; NV: Susie Lee; OH: Marcy Kaptur; PA: Madeleine Dean; SC: James Clyburn; TX: Henry Cuellar, Veronica Escobar; WA: Marie Gluesenkamp Perez; WI: Mark Pocan
Senate Appropriations Committee members:
Republicans: AK: Lisa Murkowski; AL: Katie Britt; AR: John Boozman (AR); KS: Jerry Moran; KY: Mitch McConnell; LA: John Kennedy; ME: Susan Collins; MS: Cindy Hyde-Smith; ND: John Hoeven; NE: Deb Fischer; OK: Markwayne Mullin; SC: Lindsey Graham; SD: Mike Rounds TN: Bill Hagerty; WV: Shelley Moore Capito;
Democrats: CT: Chris Murphy; DE: Chris Coons; GA: Jon Ossof; HI: Brian Schatz; IL: Richard Durbin; MD: Chris van Hollen; MI: Gary Peters; NH: Jeanne Shaheen; NM: Martin Heinrich; NY: Kirsten Gillibrand; OR: Jeff Merkley; RI: Jack Reed; WA: Patty Murray; WI: Tammy Baldwin
SOURCES
Coleman, T.W, A.D. Graves, B.W. Oblinger, R.W. Flowers, J.J. Jacobs, B.D. Moltzan, S.S. Stephens, R.J. Rabaglia. 2023. Evaluating a decade (2011–2020) of integrated forest pest management in the United States. Journal of Integrated Pest Management, (2023) 14(1): 23; 1–17
FAO. 2025. The Second Report on the State of the World’s Forest Genetic Resources. FAO Commission on Genetic Resources for Food and Agriculture Assessments, 2025. Rome.
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
results of invasion by emerald ash borer (photo courtesy of Nathan Siegert, USFS ); one of the woodboring beetles found to be so damaging to hardwood trees
Over the last nine years scientists have made significant progress in identifying aspects of insect-plant host relationships that play important roles in determining how much damage an introduced, non-native pest is likely to cause within forest ecosystems in the United States. Predicting which introductions will probably cause the greatest damage is vitally important because scientists, phytosanitary officials, and resource managers cannot address all the hundreds of established insects, much less the thousands which might be introduced. This shortfall increases with each surge in import volumes (see my previous blogs about wood packaging by scrolling down the website below the “Archives” to “Categories”, then find “SWPM”), proliferation of goods types and source areas, and cutbacks in funding.
I hope USDA APHIS and Forest Service are adjusting their procedures to apply the scientists’ path-breaking findings.
Their progress will help protect our forests. I apologize if I seem ungrateful — but we need similar progress in managing plant pathogens. Consider the damage caused by chestnut blight, white pine blister rust, Dutch elm disease, sudden oak death, beech leaf disease … (All these and other pathogens are described briefly here.) Understanding the universe of introduced fungi, water molds, nematodes, viruses, etc., is per se much more challenging. Ashley Schulz points out that among the complications are pathogens’ complex life cycles, and possible new relationships with vectors.
Undertaking this analysis will be set back decades if agencies’ resources – funds and staffs – are decimated during the current “downsizing” of government. We must speak up!!
At least regarding non-native insects that attack North American tree species, scientists’ analyses promise a new ability to set priorities. This should improve the efficacy of phytosanitary programs – if government downsizing is not allowed to destroy USDA’s scientific, regulatory, and resource management programs.
We must speak up!!
What science tells us now
Schulz et al. (2025) summarize current findings. (Full citation to all references appear at the end of the blog.)
Earlier, scientists sought to find commonalities associated with introduced insects that caused high impacts on North American conifer trees [Mech et al. (2019)] and hardwoods (angiosperms) [Schulz et al. (2021)] (Full citations at the end of the blog; earlier blogs posted here and here.) Both studies found that the time elapsed since tree species in North America diverged from the host plants of the insects in their native range (i.e., host evolutionary history) is a diagnostic factor. This factor best predicted non-native insect impact compared to the other factors that were significant for conifer and hardwood specialists. For conifers, the other significant factors included the shade and drought tolerance of the North American host plants and whether there was a related insect native to North America on the same hosts that the non-native insect impacted. For hardwoods, another important factor explaining a specialist insect’s impact is if the insect is a wood borer, especially a scolytine beetle. The wood density of the North American host plant was also considered a significant factor when predicting impact of the non-native insect.
In 2022, Uden et al. applied the divergence time method to insect species not yet introduced to North America that might attack conifer species. They hoped to identify both insects posing the greatest hazard and tree species most vulnerable to introduced pests.
Now, a new team again led by Ashley Schulz and Angela Mech (see Schulz et al. 2025) has applied a similar approach to a more comprehensive range of pest-host relationships, including the pests that specialize on host plants and pests that feed on a broader array of hosts. Some feed on both conifers and hardwoods. They found that:
It is possible to quantify insect host breadth and identify the cutoff where “specialists” and “generalists” diverge. Specifically, the split occurs around 2,250 cumulative million years, where insects that feed on hosts that add up to less than that have narrow host breadth (i.e., “specialists”) and insects that feed on hosts that add up to more than that have broad host breadth (i.e., “generalists”). This technique also helps categorize insects that fall within the middle range of host breadth and are traditionally difficult to classify as either specialists or generalists based on differing qualitative definitions of the terms.
Insects that use more hosts in their native range also tend to use more hosts in the introduced range (North America). However, many of these insects utilized fewer hosts in the introduced range compared to the native range. This shrinkage was not universal, however; about 30% of insects increased their host breadth in the introduced range. Most of these fed on a single species in their native range but attacked additional species in the same family in North America. The corresponding i-Tree Pest Predictor tool uses the list of hosts in the insect’s native range and these models to determine the insect’s likelihood that it would cause high impact, as well as each North American tree species’ susceptibility to the insect entered into the tool.
Certain feeding guilds had – on average — a significantly narrower host breadth in North America than in their native ranges. These were gall makers (13 species analyzed); sap feeders (120 species); and wood borers (35 species). In contrast, host ranges did not differ for folivores (68 species), reproductive feeders (7 species), and root feeders (5 species). Still, we know that wood borers, as a group, have caused enormous damage to a range of North American tree taxa (see emerald ash borer, redbay ambrosia beetle, invasive shot hole borers (all described briefly here). Again, the i-Tree Pest Predictor tool can help identify the threat to particular tree species.
Of course, APHIS should not disregard pests with narrow host ranges; several have caused enormous damage.
red spruce (Pikea rubens) in Great Smoky Mountains National Park; photo by famartin via Wikipedia. Red spruce is the species found to be most vulnerable by the Uden et al. study
Schulz et al. (2025) developed models for three groups of introduced herbivorous insects that feed on trees:
1) conifer specialists (based on analysis of 69 species);
2) hardwood specialists (based on analysis of 141 species);
3) hardwood generalists (based on analysis of 30 species).
Because of their quantification of host breadth, they defined the “specialist” group more broadly than is commonly done, e.g., an insect that feeds on the three families Betulaceae, Fagaceae, and Juglandaceae would be considered “specialists” because all three host families are in the Fagales clade.
Tree relatedness was the only significant explanatory factor for all three host breadth categories. As determined in the previous studies, North American host tree species that were too closely or distantly related to the insect’s hosts in its native range were less impacted than hosts that diverged somewhere in the middle – the “Goldilocks” range. The divergence period differs among the three pest-risk categories: 3–4 million years ago for conifer specialists, 5–9 million years ago for hardwood specialists, ~1–2 million years ago for hardwood generalists. Schulz et al. suggest that the reason why the peak probability of high impact differs among these groups is that different feeding guilds cause the most damage to the specific host category, and each feeding guild is challenged by different tree host defenses. Bark and wood boring beetles (the hardwood specialists with the greatest impact) must overcome lethal constitutive and induced tree defenses in order to survive for long periods in the cambial layer. These insects have adapted the ability to locate and select poorly defended individuals in the host population. Folivores (i.e., the generalists with the highest impact) adapt to plant chemistry and trichomes (hair-like or scale-like outgrowths), or can avoid host defenses by moving off the foliage. Sap feeders (which include many high impact conifer specialists) are usually tolerated by trees, unless they stimulate hypersensitive reactions or vector pathogens.
Of course, scientists’ estimates of how long ago tree taxa diverged from common ancestors differ. Fortunately, Uden et al. (2022) found that these differences only rarely affect the predicted impact of a non-native insect – at least in the case of the 62 European insects and 47 North American conifer species they analyzed. In only 1.37% of the 2,914 pairs analyzed did the predicted risk differ depending on which source phylogeny was used. These cases were associated with 27 conifer-specialist insects and 9 conifer hosts. The article does not tell us which pest/host pairs these are but, overall, this paper demonstrates that the estimate differences in the phylogenetic trees does not differ enough to be problematic when forecasting insect impact.
Changes Needed in the Way Agencies Set Priorities
Schulz et al. (2025) urged agencies to stop relying only on insect traits as the basis for developing models & phytosanitary regulations. The only insect trait that predicted impact is the insect’s feeding guild. Considering hardwoods, they found that wood borers pose the greatest risk among specialists to hardwoods; folivores among generalists. While sap feeders do not cause statistically higher damage on hardwood tree species, four of the seven high-impact conifer specialists are sap feeders (hemlock woolly adelgid, balsam woolly adelgid, red pine scale, and spruce aphid). Therefore, the i-Tree Pest Predictor tool incorporates consideration of whether a pest of conifers is a sap feeder.
Schulz et al. (2025) also caution agencies against relying on just the number of hosts an insect might exploit. Assessors must consider the range of underlying plant chemistry / host defenses that the insect encounters. They found that hosts that are shade tolerant are more susceptible to high impact from conifer specialists and hosts that have intermediate to no shade tolerance are more susceptible to high impact from generalists.
Uden et al. (2022) identified a possible weakness in USDA efforts to prioritize pest prevention targets. They found that APHIS’ Prioritized Offshore Pest List included only 12 conifer specialists from Europe among the 150 species listed. They go on to note that while sap feeders constitute 53% of tree pest species established in the U.S., APHIS listed none. The models applied by Uden, Schulz, and Mech do not consider whether the insect is likely to become established. Improving our understanding of the many factors influencing an insect’s likelihood of being transported to North America or becoming established requires additional research. This might eventually lead to a usable tool for predicting this aspect of bioinvasion by forest pests.
There is an urgent need for such a tool. As Uden et al. noted, they found that 66% of the insect species they analyzed fell into the “high impact” category. This is a much higher proportion than estimates based on earlier studies, so identifying which of these insects are likely to establish versus not establish in North America can provide more resolution and help identify which insects are going to be most problematic.
Mature Fraser fir killed by balsam woolly adelgid; Clingman’s Dome; photo by Ben Ramsey via Flickr
Tree species at risk
The analysis undertaken by Uden et al. determined that three conifer species face a high level of hazard from European insects if they are introduced. They identified particularly high threats to two species, Fraser fir (Abies fraseri) and Carolina hemlock (Tsuga caroliniana). The fir is determined to be vulnerable to 17 insect species which are predicted to have high likelihood of a high impact. The hemlock is highly vulnerable to one of the insect species they sampled. They note that both of these conifers have a limited geographic range and ecological habitat, so they likely have a relatively narrow genetic pool. A third species said to be at elevated risk is red spruce (Picea rubens) – which, although more widespread, is also under attack by a non-native insect. All three species fit earlier finding by Mech et al. that conifer trees with high shade tolerance but low drought tolerance more vulnerable to non-native pests. In none of these cases do Uden et al. mention that the tree species have already been severely diminished by established non-native insects – i.e., balsam woolly adelgid on the fir (above), hemlock woolly adelgid on the Carolina hemlock. The Schulz/Mech team is working to refine methods for identifying tree species and regions at greatest risk.
Meanwhile, Uden et al. have suggested that phytosanitary authorities and forest managers apply their findings to identify the European herbivorous insects that pose the greatest threat to North American conifer species. They should identify Palearctic tree species that fall within the high-impact “Goldilocks” zone of divergence times in relation to specific North American tree species, then identify the insects that feed on those Palearctic trees. These insects would presumably pose the highest predicted hazard to those North American tree species. They suggest that species so identified should be added to the USFS’ list of species targetted by its wood borer early detection program. To address likelihood of introduction, they suggest incorporating data on insect species commonly intercepted at ports – an indication of high propagule pressure. There will always be exceptions though. For example, Ips typographus feeds on spruce and has been frequently detected at the ports, but it has not established in North America.
For those focused on identifying species or ecoregions at greatest risk, Uden et al. suggest scientists use several sources to identify vulnerable vegetation communities. Sources suggested include USFS Forest Inventory and Analysis (FIA) and NatureServe Explorer plant community descriptions) that have relatively high-value tree species predicted to be at risk from introduced species.
SOURCES
Aukema, J.E., D.G. McCullough, B. Von Holle, A.M. Liebhold, K. Britton, and S.J. Frankel. 2010. Historical Accumulation of Nonindigenous Forest Pests in the Continental United States. BioScience 60(11): 886-897. https://doi.org/10.1525/bio.2010.60.11.5
Mech, A.M., K.A. Thomas, T.D. Marsico, D.A. Herms, C.R. Allen, M.P. Ayres, K.J. K. Gandhi, J. Gurevitch, N.P. Havill, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, A.N. Schulz, D.R. Uden, & P.C. Tobin. 2019. Evolutionary history predicts high-impact invasions by herbivorous insects. Ecology and Evolution 9(21): 12216–12230. https://doi.org/10.1002/ece3.5709
Schulz, A.N., A.M. Mech, M.P. Ayres, K. J. K. Gandhi, N.P. Havill, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, P.C. Tobin, D.R. Uden, K.A. Thomas. 2021. Predicting non-native insect impact: focusing on the trees to see the forest. Biological Invasions 23: 3921-3936. https://doi.org/10.1007/s10530-021-02621-5
Schulz, A.N., N.P. Havill, T.D. Marsico, M.P. Ayres, K.J.K. Gandhi, D.A. Herms, A.M. Hoover, R.A. Hufbauer, A.M. Liebhold, K.F. Raffa, K.A. Thomas, P.C. Tobin, D.R. Uden, A.M. Mech. 2025. What Is a Specialist? Quantifying Host Breadth Enables Impact Prediction for Invasive Herbivores
Uden, D.R., A.M. Mech, N.P. Havill, A.N. Schulz, M.P. Ayres, D.A. Herms, A.M. Hoover, K.J.K. Gandhi, R.A. Hufbauer, A.M. Liebhold, T.D. Marsico, K.F. Raffa, K.A. Thomas, P.C. Tobin, C.R. Allen. 2022. Phylogenetic risk assessment is robust for forecasting the impact of European insects on North American conifers. Ecological Applications 33(2): e2761. https://doi.org/10.1002/eap.2761
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
riparian ash killed by EAB; in this case, Mattawoman Creek in Maryland. Photo by Leslie A. Brice
Good news at the recent 33rd USDA Research Forum on Invasive Species. Scientists presented the first study that demonstrates significantly lower ash tree mortality in sites with high parasitism rates of two larval parasitoids, Tetrastichus planipennisi and Spathius galinae.
Their study area is the ash-dominated riparian area along the Connecticut River that flows north to south across the middle of Massachusetts. Knowing in advance that the emerald ash borer (Agrilus planipennis; EAB) would invade the area, scientists established monitoring plot that consisted of marked individual ash trees. EAB was first detected in the southern reach of the riparian area in 2015. It gradually moved north. By 2020 isolated mortality was observed at all sites. Meantime, they released three biocontrol agents – T. planipennis,S. galinae, and Oobius agrilii – early in the invasion at three of the six monitoring sites. These released occurred in 2018 – 2020 and again in 2022.
In 2021 and 2025, the scientists counted the numbers of biocontrol agents in the marked trees or sentinel logs. Thus the first evaluation occurred six years after EAB arrived, three years after the first releases of biocontrol agents.
They found that at southern Massachusetts sites, where EAB density was higher at the time of the biocontrol agents’ initial release, remaining ash grew more slowly than in the North. They believe the trees’ growth rate was suppressed by the trees having fewer resources. They also observed dieback. Smaller trees grew faster, perhaps responding to opening of the canopy as mature ash succumbed to EAB invasion.
The most important finding was that ash mortality at all sites was ~50% or less … not the 90% expected based on experience in the upper Midwest where the EAB invasion occurred before biocontrol agents were developed.
SOURCE
Ash survival and growth response to emerald ash borer invasion in Massachusetts riparian forests: impacts of biological control. Mitchell A. Reed, Jian Duan, Ryan S. Crandall, Roy G. van Driesche, Jeremy C. Anderson, Joseph S. Elkington. Presentation to the 33rd USDA Interagency Research Forum on Invasive Species, Annapolis, Maryland February 25-28, 2025 (The proceedings should be posted online before the end of the year.)
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
The pest alert system “PestLens” has again alerted us to plant pests in Europe or Asia that feed on species closely related to tree species native to North American forests. Two of the insects named in the alert apparently pose a hazard to icons of the forests of America’s Pacific coast forests, giant sequoia and redwood.
I hope APHIS is using this information to alert port and on-the-ground staff and perhaps initiating more in-depth risk assessments.
The posting on February 27, 2025 reported that cotton jassid, Jacobiasca lybica (Hemiptera: Cicadellidae), affects not just cotton and citrus but also Cupressus sempervirens (Mediterranean cypress) [Cupressaceae]. More than a dozen North American trees species are in this family, including
Sequoiadendron giganteum or giant sequoia. Giant sequoia is listed as an endangered species by the IUCN with fewer than 80,000 remaining in its native California.
Chamaecyparis thyoides and C. lawsoniana (Port-Orford cedar). Port-Orford cedar has been decimated in its native range by an introduced pathogen, Phytopthora lateralis. A major breeding effort has developed trees that are resistant to the pathogen; they are now available for people to plant.
Thuja occidentalis, also known as northern white-cedar, eastern white-cedar, or arborvitae,
Taxodium ascendens, also known as pond cypress
several Juniperus
Hesperocyparis macrocarpa also known as Cupressus macrocarpa, or the Monterey cypress. NatureServe ranks the cypress as GI – critically imperiled.
Cotton jassid been reported from several countries in Europe, Africa, and the Middle East.
China has reported the existence of a previously unknown bark beetle species, Phloeosinus metasequoiae (Coleoptera: Curculionidae). It was found infesting Metasequoia glyptostroboides (dawn redwood) trees in China. Affected trees exhibited reddened leaves and holes and tunnels in branches.
China has also discovered a several new hosts utilized by the fungus Pestalotiopsis lushanensis (Sordariomycetes: Amphisphaeriales). Formerly known to infect tea (Camellia sinensis) and several other plant species, P. lushanensis has now been found shoot causing blight and leaf drop on a conifer, deodar cedar (Cedrus deodara) and leaf spots on an angiospermwith congeners in North America — the rare Chinese species, Magnolia decidua. There are eight species of Magnolia native to North America.
Magnolia grandiflora; photo by DavetheMage via Wikimedia
APHIS’ ability to respond to alerts remains uncertain.
The agency’s probationary employees have been fired – just as at other agencies. APHIS staff were prohibited from participating in last week’s annual USDA Invasive Species Research Forum – the 33rd such meeting. The bird flu emergency is demanding all the attention and funds.
So – how can the rest of us fill in?
At the USDA Research Forum I again presented a poster urging greater attention to tree-killing pathogens. Scientists have made considerable progress in identifying factors that indicate whether a non-native insect might pose a significant threat (see blogs on conifer and deciduous species; more to come!). However, USDA had not funded a similar effort to improve understanding of pathogens. The most promising strategy so far are sentinel plantings. However, these systems have weaknesses; I will blog in the near future about another analysis.
I propose that APHIS start by working with independent scientists to determine the actual, current level of pathogens associated with various types of incoming goods. Contact me directly if you wish to read the text of my poster.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
emerald ash borer; some of PPA grants are funding evaluation of biocontrol efficacy
USDA APHIS has released information about its most recent annual allocation of funds under the Plant Pest and Disease Management & Disaster Prevention Program under §7721 of the Plant Protection Act. (Also see Fading Forests II and III; links provided at the end of this blog.) These funds support both critical needs and opportunities to strengthen the nation’s infrastructure for pest detection, surveillance, identification, and threat mitigation. Since 2009, this USDA program has provided nearly $940 million to more than 5,890 projects.
For FY25 APHIS allocated $62.725 million to fund 339 projects, about 58% of the proposals submitted. About $10 million has reserved for responding to pest and plant health emergencies throughout the year.
According to APHIS’ press release, the highest amount of funds (almost $16 million) is allocated to the category “Enhanced Plant Pest/Disease Survey.” Projects on “Enhanced Mitigation Capabilities” received $13.6 million. “Targetting Domestic Inspection Efforts to Vulnerable Points” received nearly $6 million. “Improving Pest Identification and Detection Technology” was funded at $5 million. Outreach & education received $4 million. I am not sure why these do not total $63 million.
Funding for States and Specific Pests
Wood-boring insects received about $2.3 million. These included more than $869,800 to assess the efficacy of biocontrol for controlling emerald ash borer (EAB) Agrilus planipennis, $687,410 was provided for various detection projects, and $450,000 for outreach efforts related to various pests. Ohio State received $93,000 to optimize traps for the detection of non-native scolytines (bark beetles).
Biocontrol efficacy will also be assessed for hemlock woolly adelgid, invasive shot hole borers, cactus moth, and several invasive plants (including Brazilian pepper). (Contact me to obtain a copy of CISP’s comments on this biocontrol program.)
Opuntia basilaris in Anza Boreggo; one of flat-padded Opuntia vulnerable to the cactus moth; photo by F.T. Campbell
Funding for other pests exceeded $1 million for spotted lanternfly (nearly $1.4 million), Asian defoliators ($1.2 million) and box tree moth (just over $1 million).
$630,000 was provided for detection surveys and studies of the sudden oak death pathogen Phytophthora ramorum, especially how it infects nursery stock. Nursery surveys are funded in Alabama, Louisiana, North Carolina, Ohio, Oklahoma, Pennsylvania, South Carolina, Tennessee, Virginia, and West Virginia. Most of these states are in regions considered most at risk to SOD infection of wildland plants.
sudden oak mortality of tanoak trees in southern Oregon; photo by Oregon Department of Forestry
Oregon received much-deserved $41,000 to evaluate the threat of the NA2 and EU2 lineages of P. ramorum to nurseries and forests Oregon also received $104,000 to respond to the detection of Phytophthora austrocedri in nurseries in the state. The Oregon outbreak has been traced to Ohio, but I see no record of funds to assist that state in determining how it was introduced.
Asian defoliator (e.g., Lymantrid moths) surveys have been funded for several years. This year’s projects are in Alaska, Arkansas, California, Kentucky, Maryland, Massachusetts, Mississippi, Montana, Nevada, North Carolina, Oregon, Tennessee, Texas, Washington, and West Virginia. While I agree that the introduction risk is not limited to coastal states with maritime ports, I don’t what criteria were applied in choosing the non-coastal states which are funded to search for these insects
Spotted lanternfly surveys (including technological improvements) or related outreach are funded in Alabama, Connecticut, Delaware, Kentucky, New Hampshire, New Jersey, North Carolina, Oregon, Pennsylvania, and Tennessee. California’s project is focused on postharvest treatments.
The Don’t Move Firewood project continues to be funded by APHIS. Several states also direct attention specifically to the firewood pathway: Kentucky, Maine, and Michigan.
I applaud the precautionary funding of the Agriculture Research Service to generate of high-quality genomic resources for managing the causal agent of Japanese oak wilt Dryadomyces quercivorous
Florida Department of Agriculture, North Carolina State University, and West Virginia University each received more than $100,000 to improve detection and management of invasive hornets.
Tennessee State University got $100,000 to continue efforts to detect and understand Vascular Streak Dieback in redbud Cercis canadensis.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
BLD symptoms; photo by Matt Borden, Bartlett Tree Experts
As beech leaf disease (BLD) is detected in an ever-expanding number of counties from Michigan to Maine south to Virginia, scientists are trying to clarify how the causal nematode — Litylenchus crenatae ssp. mccannii (Lcm) – spreads. One focus is on local spread from tree to tree. Mankanwal Goraya and colleagues set up an experiment in Stone Valley Forest, a recreation and research site managed by Penn State in Huntington County, Pennsylvania. BLD is present – although I have not been able to determine for how many years. [The full citation to Goraya et al. is provided at the end of this blog.]
Goraya et al. (2024) set up four stands, each bearing three funnels, at varying distances from naturally BLD-infected American beech (Fagus grandifolia) trees. Two stands were at 3.51 m from symptomatic trees of starkly different sizes: one of the trees had a dbh of 50 cm, the other of only 5.6 cm. A third close-up stand was set up at 2.20 m from another large tree, having a dbh of 46 cm. The fourth stand was set up at a significantly longer distance, 11.74 m from a symptomatic beech tree; this tree was also small, with a dbh of 5 cm. This arrangement allowed the scientists to detect influences of both distance from the source of infection and relative canopy size of the source tree. They consider dbh to be an adequate substitute for canopy size. There was apparently no other effort to determine or vary the height of “source” trees, although I think that might influence speed of the wind flowing through the canopy.
Goraya et al. also tested whether it is possible to detect the presence of Lcm in association with other invertebrates that live in beech forests. To do this, they counted numbers of nematodes in frass from six species of caterpillars that had been feeding on leaves of infected trees, and in two spider webs spun in the branches of symptomatic trees. They also determined whether these nematodes were alive (active) or inactive – presumably dead.
The study makes clear that Lcm’s life cycle and impact are not as surprising as initially thought. Several species in the family Anguinidae – to which Lcm belongs – are considered significant pests. These nematodes can parasitize aerial parts of the plants (leaves, stems, inflorescences and seeds), causing swellings and galls. Furthermore, they are migratory; they can move across the surface of host tissues using water films. Once they have penetrated the host tissues, they can induce host cell hyperplasia and hypertrophy, resulting in leaf or bulb deformities, shorter internodes, and neoplastic tissues. Furthermore, heavy rainfall and wind are known to play significant roles in the dissemination of plant-infecting nematodes. In their desiccated state on infected seeds, some species of this family can survive passage through animals’ gastrointestinal digestive tract (e.g., domestic livestock, insects, & birds).
A crucial factor is that Lcm can reach densities of thousands of nematodes per leaf by late summer or early fall, increasing the likelihood of their exposure to facilitating environmental conditions at the time they migrate from leaves to buds. And once established within the bud tissues, the nematodes feed on bud scales and newly forming leaves to develop & increase their pop #s. They also use the bud as protection from adverse environmental conditions.
Goraya and colleagues collected samples every other day from September 9 to November 23, 2023 – the period when Lcm migrate from highly infected leaves to newly forming buds. [I note that it in the mid-Atlantic – where Lcm is spreading – we had an extensive drought in autumn 2024 – more than 30 days without any rain from early October into November. I hope scientists are monitoring BLD spread sufficient closely to see whether this drought affected dispersal.]
Nematodes dispersal linked to weather
Goraya and colleagues collected 324 samples from the funnels. Eighty-two percent (n =266) of the samples had nematodes; up to 92% were identified as Lcm. Non-Lcm nematodes were distributed across different genera, mostly classified as free-living nematodes. While several hundred nematodes were found in the funnels on most days, numbers peaked noticeably on some days in September and October. A startling 2,452 nematodes were recovered from a single funnel in October. Depending on the sample, up to 67% of Lcm recovered from the funnels were active.
Analysis of the environmental (weather) variables found that increases in wind speed, humidity, and precipitation (rainfall) coincided with higher numbers of Lcm being recovered from the funnels. However, the effect of wind speed becomes less positive as precipitation increases or vice versa. Goraya et al. suggest a pronounced negative interaction between wind and rain. At low precipitation levels, increased wind speed might facilitate Lcm dispersal. As rainfall increases, higher wind speeds might carry the Lcm nematodes farther away. Support is seen in the fact that fewer nematodes were found in the funnels closer to the BLD-infected trees during these periods. Really heavy rain might push a significant preponderance of nematodes to the ground. The scientists point to a very complex interplay between weather patterns and Lcm population dynamics and dispersal.
BLD symptoms on beech tree in Fairfax County, Virginia – a dozen miles from known infestation; photo by F.T. Campbell
The model did not show any significant influence of maximum temperature on nematode numbers in autumn. Goraya et al. do not speculate on whether temperatures might play a role during summer, as distinct from cooler autumn periods.
Goraya et al.’s findings differ from those of previous studies. Earlier documentation of wind dispersal of nematodes concerned primarily free-living species. It was unexpected to find consistently much higher numbers of Lcm – especially because Lcm is a plant-parasitic nematode. Another surprise is the high proportion of nematodes that are active.
Goraya et al. conclude that because Lcm is actively migrating in large numbers during autumn months, it is primed to take advantage of favorable weather. This nematode will likely survive and thrive in the environmental conditions of beech forests in northeastern North America.
Considering the effect of distance, some findings fit expectations: significantly more Lcm were recovered from funnels placed near symptomatic “source” trees than from those farther away. However, this was not a simple relationship. For example, in two cases the scenarios seemed nearly alike: both “source” trees were large (dbh 46 or 50 cm) and symptoms were “medium-high” (more than half of leaves presenting dark-green interveinal bands). Distance of funnels from the “source” tree differed minimally: 2.2 m versus 3.51 m. Still, the number of nematodes retrieved from the two sets of funnels differed significantly: one set of funnels recovered the highest number of Lcm nematodes obtained during the entire experiment – 2,452; the second contained only up to 600 nematodes. The authors do not offer an explanation.
I am not surprised by the apparently strong correlation between numbers and proximity to the disease source (a symptomatic tree). Nor am I surprised that Lcm nematodes were also found in funnels 11 meters away. I do wonder, however, why they are certain that no source was closer. Detecting early stage infections is notoriously difficult.
beech with large canopy; photo by F.T. Campbell
Goraya et al. also evaluated the effect of size of the source tree. They used dbh a substitute for larger canopies. Trees with larger canopies can host more nematodes, so are likely to contribute more to dispersal events. Two sets of funnels were equidistant from separate “source” trees – 3.51 m. One tree was small – 5.6 cm dbh, 11% as large as the other tree (50 cm). They collected many fewer Lcm nematodes from the smaller tree – the maximum was only 132 compared to 600 (a decrease of 78%).
Still, small trees can apparently support spread of the nematode to a reasonable distance. The fourth set of funnels was set up more than three times farther away (11.74 m) from an infected tree of a similar size (dbh = 5 cm) but recovered almost the same number of Lcm nematodes (0 – 119).
I find it alarming that both small trees in this part of the experiment had low BLD symptoms – only a few leaves were banded. Yet they apparently are the source of Lcm spread. The alternative, as I noted above, is that other “source” trees were in the vicinity but were not detected, possibly because they did not yet display symptoms?
Goraya et al. conclude that “source” tree size directly impacts the number of recovered nematodes. In addition, wind plays a pivotal role in their local distribution. This suggests a complex dispersal pattern in which proximity to the source leads to higher numbers of nematodes but longer-distance spread is possible.
Tussock moth; photo by Jon Yuschock via Bugwood
Nematodes’ association with other organisms
Goraya et al. (2024) collected one each of six caterpillar species from BLD-symptomatic trees. The frass of one – the tussock moth caterpillar (Halysidota tessellaris) — contained 12 nematode specimens — 10 of them Lcm. Two of the Lcm were alive and active. Their presence indicates that Lcm can survive passage through the caterpillar’s gastrointestinal tract. The authors conclude that caterpillars feeding on symptomatic leaves might contribute to local dispersal of Lcm.
Hundreds of Lcm were recovered from the two spider webs collected from the branches of a BLD-infected beech tree. From one web, 255 nematodes were captured; 58 were active. In the second web there were only 34 Lcm, but one-third — 10 – were active.
Goraya et al. (2024) hypothesized that any biotic form having the ability to move from a BLD-infected tree would be able to transport Lcm to other non-infected trees. Beyond caterpillars, they speculate that birds consuming these caterpillars might also disperse Lcm. Doug Tallamy has documented that many birds feed on caterpillars, link although he is focused on those that consume caterpillars in the spring, not the autumn. They note that others are studying that the bird species that feed on beech buds (e.g., finches) might transport nematodes. They note the need for additional research to clarify whether the nematode can survive birds’ digestive system.
Re: detection of live Lcm in spider webs, Goraya et al. suggest two possible interpretations: 1) this finding demonstrates that nematodes might fall from leaves, potentially spreading the infection to other trees beneath the canopy. (Supporting this idea is the fact that sub-canopy trees are often heavily infected with BLD and are frequently the first to exhibit BLD symptoms.) 2) Nematodes in spider webs are very likely to be transported by other “incidental organisms” (e.g., insects, birds, mammals) that feed on invertebrates trapped in webs — thereby potentially increasing the number and impact of nonspecific nematode vectors.
In conclusion, Goraya et al. found that many factors, e.g., distance & size of infected beech trees, wind speed, & humidity, contribute significantly to Lcm dispersal. The multitude of organisms interacting beneath the canopy also play a role.
They suggest that several major questions still need to be explored. These include how Lcm navigate environmental factors in their spread; and whether Lcm can survive – perhaps in a anhydrobioses state –transport over long distances, whether by abiotic or biotic vectors.
I remind my readers of the importance of beech in the hardwood forests in northeastern North America. Many wild animals, including squirrels, wild turkeys, white-tailed deer, and bears depend on beechnuts for fats and proteins. Moreover, some insects birds rely on beech tree canopies for shelter & nesting.
Other Hosts
Beech leaf disease attacks not just American beech (Fagus grandifolia). In North America, it has also attacked planted European beech(F. sylvatica), Chinese beech (F. engleriana), and Oriental beech (F. orientalis). Thus if it spreads it could have severe impacts across forests of much of the Northern Hemisphere.
range of European beech; from Royal Botanic Gardens, Kew
I appreciate that this project was funded by the USDA Forest Service International Program. I will pursue information concerning efforts by USFS Research and Development and the Forest Health Protection program.
SOURCE
Goraya, M., C. Kantor, P. Vieira, D. Martin, M. Kantor. 2024 Deciphering the vectors: Unveiling the local dispersal of Litylenchus crenatae ssp mccanni in the American beech (Fagus grandifolia) forest ecosystem PLOS ONE |https://doi.org/10.1371/journal.pone.0311830 November 8, 2024 1 / 16
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/