red deer on farm in New Zealand; photo by Bernard Spragg via Flickr
Among the non-native species damaging forest systems are mammals – introduced deer, goats and sheep, and swine, … These animals have the greatest impacts on island systems that are sufficiently isolated that they have no native terrestrial mammals, e.g., Hawai`i and New Zealand. Several New Zealanders have published a study of their impacts (Allen et al.; full citation at end of the blog). The focus of their analysis is the native forests’ ability to sequester carbon and thus mitigate climate change. The scientists are well aware, however, that forests provide many other ecosystem values and services, including biodiversity, water supply and quality, etc.
Introduced ungulates can have many direct effects: reduction and damage to understory biomass, depletion of seedling regeneration, exacerbated soil erosion, and local nutrient imbalances. Mammals’ browsing can modify the composition of plant communities by favoring abundance of unpalatable species. Changes also can alter ecosystem functions associated with nutrient cycling, e.g., by reducing nutrient returns to the soil and altering rates of litter decomposition
In these ways, introduced ungulates exert long-term impacts on forests’ capacity to store carbon.
Allen et al. aimed to determine the extent of these effects on forests’ capacity to store carbon, both above- and below-ground, and on forest structure and diversity. The authors compared data from 26 pairs of sites across New Zealand – half with ungulate exclosures and half adjacent unfenced control plots. The ungulate exclosures had all been established for at least 20 years. All the sites were in species-rich communities of conifers and broadleaved evergreen angiosperm trees. These forests (1) cover about one-third of the country’s remaining mature natural forest; (2) contain tree species of a wide range of palatability to ungulate herbivores; and (3) have been named a conservation priority for forest carbon management. The ungulates present on the plots were European red deer (Cervus elaphus), fallow deer (Dama dama), sika deer (Cervus nippon), and feral goats (Capra hircus).
They assert that New Zealand is a good place to do this type of study because ungulate introductions are relatively recent so their impacts are well documented.
Allen et al. found that managing invasive ungulates makes valuable contributions to conserving biodiversity but not to carbon sequestration. They found little difference in total ecosystem carbon between ungulate exclosures and unfenced control plots. Most of the difference they did find was explained by the biomass of the largest tree within each plot. As they point out, these large trees have been unaffected by invasive ungulates introduced during the last 20–50 years. However, they believe ungulate-caused changes in understory biomass, species composition, and functional diversity might result in major shifts in the diversity and composition of regenerating species. Hence, longer term consequences for both ecosystem processes and storage of forest carbon storage can be expected.
Indeed, excluding ungulates did increase the abundance and diversity of saplings and small trees. The basal area of the smallest class of tree size was 70% greater. Species richness of small trees and saplings was 44% and 68% higher, respectively. This difference had little impact on overall carbon storage, however, because the small trees and saplings store only about 5%. In contrast, the largest tree size class (dbh =/>30 cm), with their roots, contributed 44% of total ecosystem carbon in both exclosure and control plots. The largest effects of exclosures on carbon stocks were in early successional stands, e.g., those affected by such major disturbances as windthrow, volcanic activity, or landslides.
Climate change is expected to cause surprising interactions among forest productivity, herbivory, disturbance. Allen et al. suggest that authorities should focus on excluding ungulates on these highly productive regenerating forests rather than old-growth forests. I am disturbed by this suggestion. It exposes the most biologically diverse forests to continuing damage.
Data gaps
New Zealand has many long-lived, slow-growing tree species. Recruitment of understory trees is already low across both main islands. This situation has been attributed to ungulate browsing. Over centuries, this might result in shifts in the canopy composition. Allen et al. call for additional research to increase our understanding of how browsing and other short-and long-term drivers affect the regeneration of large trees. Also, data on soil CO2 emissions needs better integration.
Australian brushtail possum; photo by Peter Firminger via Flickr
The study did not consider the impact of other introduced mammals, such as feral pigs (Sus scrofa), rodents, and Australian brushtail possum (Trichosurus vulpecula). The possum is known to damage New Zealand trees. The scientists did not explain this omission; I assume it might have been the result of either lack of resources to support a broader study or differences in management strategies – or both?
I note that the study also did not address the extent to which non-native pathogens threaten these large trees. In response to my query, Kara Allen said that their plots did not include many kauri (Agathis australis) trees, so the severe dieback disease caused by Phytophthora agathidicida did not affect their results. Naturally regenerating kauri is limited to a small area of warm temperate rainforests located at the top of the North Island. So kauri potentially play a relatively small role in terms of overall carbon stocks in New Zealand’s forests. On the other hand, Allen says thatmyrtle rust (Austropuccinia psidii) could have a major impact on New Zealand forests’ carbon storage. Trees in the host family, Myrtaceae, are ecologically important across both islands. Also, they comprise a large portion of overall forest carbon stocks (ranked in the top 5 largest families for above- and belowground biomass). An example is southern rata (Mterosideros umbellata), which are preferentially fed on by Australian brush possum.
southern rata; photo by Stan Shebs
Bernd Blossey, (free access!) who has long studied the role of high deer populations in North American forests, praises the study’s attempt to measure data, not just rely on models, and its inclusion of soil. However, he notes other limitations of the New Zealand study:
The small exclosures (20 x 20 m) are subject to edge effects. Some of Blossey’s exclosures occupy 2 hectares.
Twenty years is too short a time for analysis of such long-term processes as carbon sequestration and regeneration of slow-growing trees. Therefore, any results must be considered preliminary. Furthermore, no one recorded any differences in carbon sequestration of the paired plots at the time the exclosures were set up.
There’s no mention of possible impacts by introduced earthworms.
Dr. Blossey recognizes that the current study’s authors cannot re-do actions taken decades in the past. Still, the data gaps reduce the value of the findings.
I conclude that uncertainties continue due to: the long timelines of species’ regeneration and growth to full sizes; the requirement for large exclosures; the complexity of factors affecting carbon sequestration; and probably other influences.. Managers trying to maximize carbon sequestration are forced to act without truly knowing the best strategy or how their actions will affect the future.
For more about invasive mammals’ impacts in U.S. forests, also see the study by USFS scientists, Poland et al. (full citation listed in sources). One can enter “mammal” in the search box for the on-line PDF.
SOURCES
Allen, K., P.J. Bellingham, S.J. Richardson, R.B. Allen, L.E. Burrows, F.E. Carswell, S.W.Husheer, M.G. St. John, D.A. Peltzer, M. Whenua. 2023. Long-term exclusion of invasive ungulates alters tree recruitment and functional traits but not total forest carbon. Ecological Applications. 2023; e2836. https://onlinelibrary.wiley.com/r/eap
Poland, T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019), Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag. The on-line version as at https://link.springer.com/book/10.1007/978-3-030-45367-1
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
spread of non-native conifers in mountains of New Zealand; photos by Richard Bowman; New Zealand government website
As those of us who want to “do something” to counter bioinvasions struggle to mobilize both the resources and the political will necessary, I rejoice that more studies are examining what factors affect “social license” [= public approva] for such programs. One such study was recently published in New Zealand — Mason et al. (full citation at the end of the blog). New Zealand enjoys a greater appreciation of the uniqueness of its biology and awareness of invasive species’ impacts than the United States. However, their findings might provide useful guidance in the US and elsewhere.
Mason et al. sought to understand motivations of, and constraints on, those local groups responsible for controlling the spread of non-native conifers into New Zealand’s remnant native ecosystems. Non-forest ecosystems across much of the country are at risk of rapidly transforming into exotic conifer forests. For these reasons, authorities are pressing for timely removal of existing seed sources, that is, mature non-native conifer trees of several species. The blog I posted earlier apparently describes effects of conifer invasions in lowland ecosystems, whereas the Programme described here is focused on high-elevation systems.
The eradication effort in the study is the National Wilding Conifer Control Programme, establishedin 2016. A large increase in funding provided during the COVID-19 lockdown made it practical to try to eradicate seed sources from large swathes of vulnerable land. The Programme coordinates control efforts across the country, working across property and land-tenure boundaries. Landowners are expected to cover 20% of the cost of removing conifers from their land. Since removing all seed sources of high-risk conifer species from the landscape is key to achieving long-term goals, success is unlikely if significant seed sources are allowed to persist.
Mason et al. combined workshops, questionnaires, and site visits to gather data on particular aspects of this Programme. They found that social resistance, rather than lack of scientific knowledge, was often the main barrier to success in managing widespread invasive species. The authors do not address whether the fact that only 30 people provided information for their study might undermine the reliability of their findings.
map of conifer wilding sites; adapted from Wilding conifers – New Zealand history and research background, a presentation by Nick Ledgard at the “Managing wilding conifers in New Zealand – present and future” workshop (2003)
The authors suggest that the main benefit of scientific information might be to increase stakeholders’ support for management interventions — rather than to guide manager’ decisions about which strategies to pursue. To support social license, invasive species research programs might need to focus not only on cost-effective control technologies and strategies, but – perhaps especially — the benefits (both tangible and intangible) of invasive species control for society.
Mason et al. found that people were motivated to combat conifer invasions by impacts with direct influence on humans or human activities (e.g., reduced water yield, damage to infrastructure from wildfires, reduced tourist activities due to landscape transformation) and also by impacts affect ecosystems (e.g., impacts on biodiversity, aquatic ecosystems and landscapes).
People objected to control or eradication programs primarily because of social concerns. These included the unwillingness of landowners to participate and regulatory frameworks that had perverse incentives.
Mason et al. called for greater efforts by scientists to persuade stakeholders[p1] to allow removal of “wilding” conifers from private land and development of more appropriate regulations. They found that forecasting models were particularly effective in persuading people to support these efforts. It seems to me that outreach teams might need “translators” to convert scientists’ findings to information that would be more useful by stakeholders.
The authors concede that the “wilding conifer” situation has unique attributes. First, invading conifers present a stark, easily seen difference between native and invaded ecosystems. Second, some – but not all—stakeholders appreciate the uniqueness of New Zealand’s biomes. Third, the impacts of conifer invasion are sufficiently well known that they can be described succinctly and accurately.
Do these unique attributes undercut the relevance of this study to North America? It is still true that ongoing support from local stakeholders (including landowners and community groups) influences the effectiveness or profitability of managing invasive species. .It is also true that groups’ varying values affect willingness to support the activities.
Mason et al. think through the issue of stakeholders’ conflicting perspectives on the value of particular invasive species and the values threatened by that invader. These can include ethical or safety concerns around management methods, particularly regarding toxins and genetic modification. Economoic costs are also a factor – especially if the landowner must pay all or some of them.
I find it interesting that the government simultaneously funded a 5-year research program to study various issues regarding the spread, ecosystem impacts, and control of wilding conifers. The result is the Mason et al. study discussed here. I wish the U.S. funded independent analyses of its invasive species programs!
Pinus contorta – the most rapidly growing Pinus introduced to New Zealand; photo by Walter Siegmund / Wikimedia
More Details, Policy Suggestions
Workshop attendees unanimously identified landscape impacts as a reason for controlling wilding conifers. This primarily concerned the loss of New Zealand’s visual heritage or cultural identity rather than loss of native species’ habitats. When the landowner was raised in Europe, these cultural or heritage values sometimes had the opposite effect, since they see conifer forests as important components of “natural” landscapes.
Currently, landowner funding and permission is required for conifer removal. Some individual landowners want to establish new forestry plantings. Some resist removal of existing forestry plantations (which provide income) and shelter belts (which provide shelter for livestock in high country landscapes). Some landowners were unwilling to pay their 20% of removal costs. Or they objected to certain conifer control methods—particularly helicopter spraying of herbicides. New Zealand’s regulatory process also requires years of negotiating to remove standing trees – further delaying any action. In theory, landowners who resist removal could be prosecuted under the Biosecurity Act. However, this approach has never been tried for removing wilding conifers.
Mason et al. suggested several changes in policy to overcome some of these barriers.
First, forestry consultants can “game” the wilding conifer “risk calculator” to obtain government approval to establish conifer plantations in high-risk environments. The authors suggest that authorities create a “liability calculator.” Under this system, landowners wishing to retain conifers on their land for whatever reason would be liable for any subsequent containment costs. However, developing such a tool requires more finely-scaled models of conifer spread.
Second, given the high costs of combatting invading conifers if seed sources are allowed to persist, they suggested that it might be more cost-effective for the control program to pay for plantation removal under New Zealand’s Emissions Trading Scheme.
Given the overwhelmingly social and regulatory nature of barriers to success, the primary role for scientific information is providing assessments of outcomes in the absence of wilding conifer control. Preferred messages were return-on-investment estimates and forecasts of ecosystem impacts, particularly relating to biodiversity loss, water yield reduction, and wildfire hazard. Forecasts were key to demonstrating that management interventions reduced future control costs and avoided environmental impacts which large sections of the community value (i.e. biodiversity loss, reduction in water yield and agricultural productivity, increased wildfire risks). Practitioners felt that forecasting models might also channel research toward areas of high uncertainty. Mason et al. recognize the difficulties presented by inherent complexity of ecological systems. However, they think “good practice” guidelines on forecasting are emerging.
The authors find that information content and presentation need to be tailored to the various audiences – most of whom lack experience in interpreting data from environmental forecasting models. They suggest that outreach materials focus on clear illustration of the tangible and intangible benefits of wilding conifer management rather than detailed explorations of scenarios. Participants suggested ways to improve the web tool to make it more accessible to a non-expert audience.
Mason et al. mention aspects that require balancing, but don’t suggest criteria for making these choices. They say it is important to include all relevant stakeholders in invasive species management governance bodies. The absence of stakeholders with positive attitudes to wilding conifer invasions led to unanticipated external social resistance to the Programme. They recognize that including stakeholders with conflicting interests might obstruct the decision-making process. Also, in areas where there has been success in containing conifers’ spread, people can’t see invading trees, so they don’t recognize the problem. They also note that existing data do not adequately recognize risks of spread from deliberately planted seed sources such as shelter-belts, plantations and amenity plantings. The authors do not discuss how to integrate these data into analyses and public outreach.
Finally, Mason et al. recognize that many other factors strongly influence stakeholders’ willingness to support invasive species control programs, especially the level of trust and strength of relationships between bioinvasion program staff and stakeholders.
Also, they suggest topics for future research: assessing how well forecasting models are integrated with communications with stakeholders; how qualitative and quantitative research methods in different fields might support one another; and empirical tests to measure the relative effects on social license of a) involving stakeholders in developing models, b) using forecasts to assess the consequences of different management decisions and, c) the usefulness of different methods for incorporating scientific information in stakeholder engagement.
SOURCE
Mason, N.W.H., Kirk, N.A., Price, R.J. et al. Science for social license to arrest an ecosystem-transforming invasion. Biol Invasions25, 873–888 (2023). https://doi.org/10.1007/s10530-022-02953-w
see also https://www.doc.govt.nz/nature/pests-and-threats/weeds/common-weeds/wilding-conifers/
Posted by Faith Campbell
What do YOU think about the role “social license” plays in US invasive species programs? We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
This blog asks YOU!!! to support funding for some of the key USDA programs. This blog focuses on USDA’s Animal and Plant Health Inspection Service (APHIS). APHIS is responsible for preventing introduction of pests that harm agriculture, including forests; and for immediate efforts to eradicate or contain those pests that do enter. While most port inspections are carried out by the Department of Homeland Security Bureau of Customs and Border Protection, APHIS sets the policy guidance. APHIS also inspects imports of living plants.
Please help by contacting your members of the House and Senate Appropriations Committees. I provide a list of members – by state – at the end of this blog. APHIS is funded by the House and Senate Appropriations Subcommittees on Agriculture and Related Agencies. These Subcommittees have scheduled hearings on the topic and I’ve drafted written testimony for them. I expect CISP will be joined by additional members of the Sustainable Urban Forest Coalition in signing the testimony. You can add the crucial voice of constituent’s support.
I will blog soon about funding for USDA’s Forest Service (USFS) – I don’t yet have necessary information to suggest specific funding levels.
Your letter or email need be no more than a couple paragraphs. To make the case for greater funding, feel free to pick-and-choose from the information that follows. Your greatest impact comes from speaking specifically about what you know and where you live.
These are the specific dollar amounts we’d like you to ask for. The rationale for each is below.
Appropriations for APHIS programs (in $ millions)
Program
FY 2022 (millions)
FY 2023
FY 2024 Pres.’ request
Our ask
Tree & Wood Pest
$61
$63
$64
$65 M
Specialty Crops
$210
$216
$222
$222 M
Pest Detection
$28
$29
$30
$30 M
Methods Development
$21
$23
$23
$25 M
The Costs of Introduced Pests
Introduced pests threaten many forest products and services benefitting all Americans, including wood products, wildlife habitat, carbon sequestration, clean water and air, storm water management, lower energy costs, improved health, aesthetic enjoyment, and related jobs. Already, the 15 most damaging non-native pests threaten at least 41% of forest biomass in the “lower 48” states. In total, these 15 species have caused an additional annual conversion of live biomass to dead wood at a rate similar in magnitude to that attributed to fire (5.53 TgC per year for pests versus 5.4 to 14.2 TgC per year for fire) [Fei et al.; full citation at end of blog; see also earlier].
tanoaks killed by SOD; Oregon Department of Forestry photo
These pests also impose significant costs that are borne principally by municipal governments and homeowners. As more pests have been accidentally introduced over time, these costs have risen. A study published last year [Hudgins et al.] projected that by 2050 1.4 million street trees in urban areas and communities will be killed by introduced insect pests. Municipalities on the forefront include Milwaukee and Madison Wisconsin; the Chicago area; Cleveland; and Baltimore, Towson, and Salisbury, Maryland. Removing and replacing these trees is projected to cost cities $30 million per year. Additional urban trees – in parks, on homeowners’ properties, and in urban woodlands – are also expected to die and require removal and replacement.
Pathways of Introduction
Tree-killing pests are linked to the international supply chain. Many pests—especially the highly damaging wood-borers like emerald ash borer, Asian longhorned beetle, polyphagous and Kuroshio shot hole borers, and redbay ambrosia beetle—arrive in inadequately treated crates, pallets, and other forms of packaging made of wood. Other pests—especially plant diseases like sudden oak death and sap sucking insects like hemlock woolly adelgid—come on imported plants. Some pests take shelter, or lay their eggs, in or on virtually any exposed hard surface, such as steel, decorative stone, or shipping containers.
infested wood from a crate; Oregon Department of Agriculture photo
Wood Packaging
Imports from Asia have historically transported the most damaging pests, e.g., Asian longhorned beetle, emerald ash borer, redbay ambrosia beetle, and the invasive shot hole borers. For decades goods from Asia have dominated imports. As of February 2022, U.S. imports from Asia were running at a rate of 20 million shipping containers per year. A recent analysis [Haack et al.; see also here] indicates that at least 33,000 of these shipping containers, perhaps twice that number, are carrying a tree-killing pest. These facts have led scientists to project [Leung et al.] that by 2050, the number of non-native wood-boring insects established in the US could triple. Hudgins et al. say the greatest damage would occur if an Asian wood-boring insect that attacks maples or oaks were introduced. Such a pest could kill 6.1 million trees and cost American cities $4.9 billion over 30 years. The risk would be highest if this pest were introduced to the South – and U.S. southern ports are receiving more direct shipments from Asia after the expansion of the Panama Canal in 2016. https://www.nivemnic.us/?m=202207
After introduction of the ALB, APHIS acted to curtail further introductions in wood packaging from China. First – in 1998 – APHIS required China to treat its wood packaging. Second, it worked with foreign governments to develop the International Standard for Phytosanitary Measures (ISPM) #15. The U.S. and Canada began phasing in ISPM#15 in 2005 with full implementation in 2006. Under ISPM#15, all countries shipping goods to North America must treat their wood packaging according to specified protocols with the goal of “significantly reducing” the risk that pests will be present.
However, as I have often blogged [see blogs under “wood packaging” category on this site] ISPM#15 has fallen short. Haack et al. found that as recently as 2020, 0.22% [1/5th of 1 percent] of the shipping containers entering the U.S. were infested by a tree-killing insect. This equates to tens of thousands of containers harboring tree-killing insects.
Worse, the data indicate that our trade partners’ compliance with the rules has deteriorated; the “approach rate” of pest-infested wood packaging fell in 2005-2006, but has since gone back up.
The most troubling offender is China. Although required since 1998 to treat its wood packaging, China consistently has one of the highest pest approach rates: it was 0.73% [or ¾ of 1%] during the 2010-2020 period. This is three times the global average for the period. Since China supplied 40.7% of U.S. imports in 2022 [Szakonyi], or 5,655,000 containers. Thus China alone might be sending to the U.S. 30,000 containers infested with tree-killing insects. These pests threaten our urban, rural, and wildland forests and reduce forest productivity, carbon sequestration, the rural job base, water supplies and quality, and many other ecosystem services.
ISPM#15 falls short at the global level. The fact that a pallet or crate bears the mark indicating that it complies with ISPM#15 has not proved to be reliable.
You might ask your Member of Congress or Senators to ask APHIS what steps it will take to correct the problem of Chinese non-compliance. (Remind him or her that that the Asian longhorned beetle, emerald ash borer, and many other insects of so-far lesser impact were introduced in wood packaging from China.
Asian longhorned beetle
Remind them also that the Department of Homeland Security’s Bureau of Customs and Border Protection has twice enhanced its enforcement of wood packaging rules. In 2017 it began penalizing importers of non-compliant wood packaging under Title 19 United States Code (USC) §1595a(b) or under 19 USC §1592. In 2021, it incorporated the wood packaging requirements into its voluntary C-TPAC program.)
You might also urge them to ask APHIS what steps it is taking at the global level to improve the efficacy of ISPM#15 – or to replace it if necessary to ensure that pests are not being introduced.
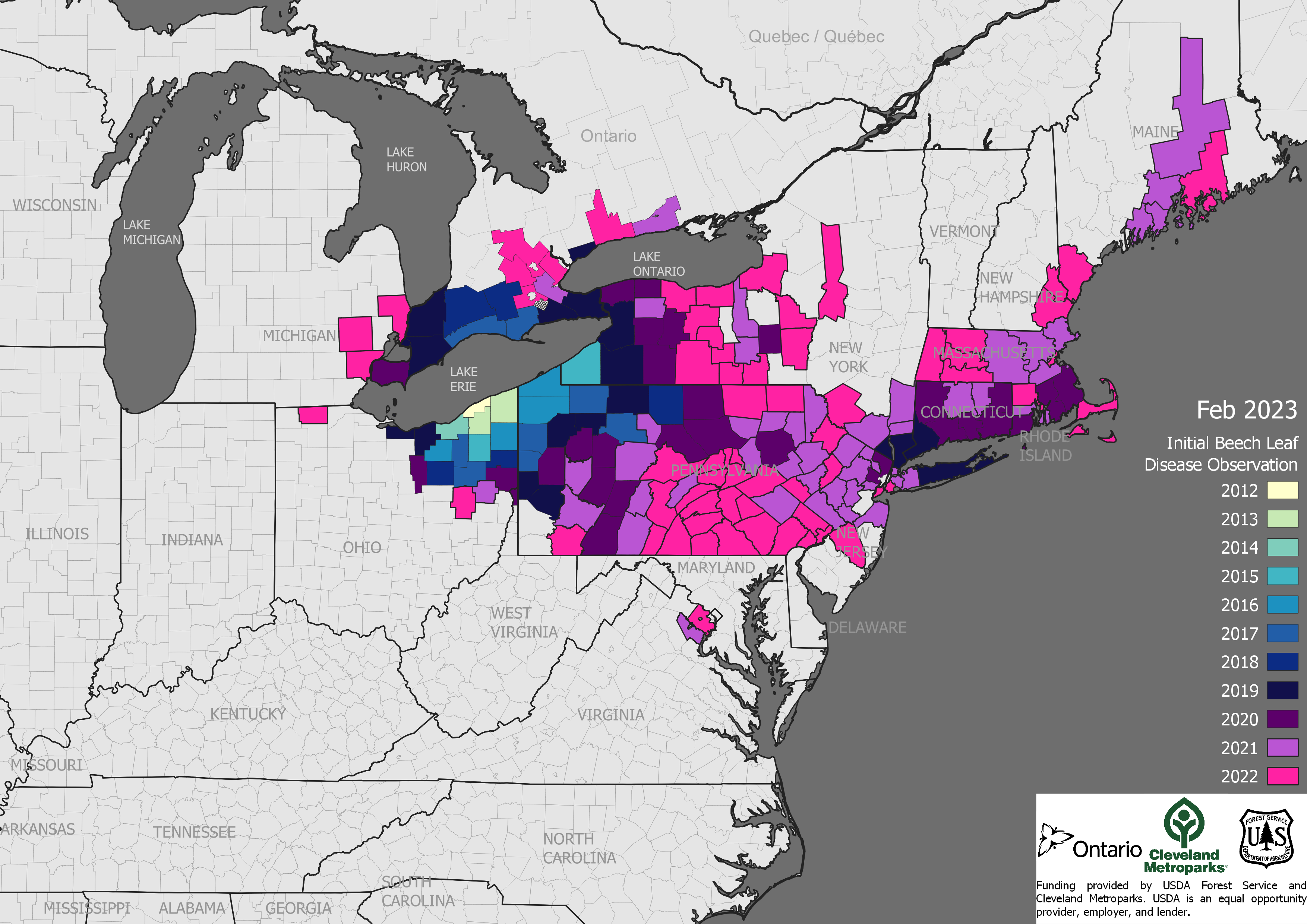
spread of beech leaf disease
Imported Plants (“Plants for Planting”)
Some pest types—especially plant diseases like sudden oak death and sap-sucking insects like hemlock woolly adelgid—come on imported plants. The U.S. imported about 5 billion plants in 2021 [MacLachlan]. Recent introductions probably via this pathway include several pathogens — Phytophthoras, rapid ʻōhiʻa death in Hawai`i, beech leaf disease (established from Ohio to Maine), and boxwood blight. Insects have also been introduced on imported plants recently; one example is the elm zigzag sawfly (present in North Carolina, Virginia, and New York and Ontario). https://www.nivemnic.us/?p=4115
An analysis of data from 2009 [Liebhold et al.] found that approximately 12% of plant shipments were infested by a pest. This pest approach rate is more than 50 times higher than the 0.22% approach rate for wood packaging. APHIS has adopted several changes to its phytosanitary system for imported plants in the decade since 2009. A few studies have been published, but they have focussed on insects and excluded pathogens. We have noted that pathogens continue to be introduced via the plant trade. Therefore, please ask your Member or Senators to ask APHIS to facilitate an independent analysis of the efficacy of the agency’s current phytosanitary programs to prevent introductions of pests on important plants, with an emphasis on introductions of plant pathogens.
APHIS is responsible for preventing spread of the SOD pathogen, Phytophthora ramorum, through trade in nursery plants. In recent years California has had few detections in nurseries and little expansion in forests – but the situation suggests that this good news is probably more the result of the drought than of program efficacy. In cooler, wetter conditions in Oregon and Washington, detections in nurseries and alarming detections in the forest or plantings continue.
In 2022, the APHIS SOD Program supported detection and regulatory activities in 25 states. P. ramorum was detected at 18 establishment, 12 of which were first-time detections. The California nursery regulatory program – which is funded by APHIS – saw reduced funding in 2022. We think these cuts are unwise since this year’s very wet winter will probably lead to a new disease outbreaks. Programs in Oregon and Washington continue to detect infestations in additional retailers brought in by plants bought from other nurseries. Washington responded to four separate “trace forward” incidents, one involving more than 160 residential sites. Clearly, the federal-state program is not succeeding in eradicating P. ramorum from nurseries. Please suggest that your Congressperson and Senators ask APHIS what steps it is taking to improve the efficacy of the SOD program.
SOD-infected rhodoendron on plants in Indiana; photo by Indiana Department of Natural Resources
In the East, P. ramorum was found in three of 65 streams sampled in 10 states in 2022 (reaching across the Southeast from Mississippi through North Carolina, plus Texas, Maryland, Pennsylvania, and Illinois). One stream is troubling: a first-time detection in South Carolina, with no obvious nursery source. Since stream sampling began, P. ramorum has been detected from eight streams in four states, Alabama, Mississippi, North Carolina, and now South Carolina. The pathogen has been present in some of these streams for more than 10 years.
Oregon faces particularly high risks. Three of the four known strains of P. ramorum are established in Oregon forests. One of them, the EU1 lineage, is more aggressive than the NA1 clonal lineage already present in forests. In addition, the EU1 strain might facilitate sexual reproduction of the pathogen, thus exacerbating Oregon’s struggle to contain the disease.
As we know, introduced pests do not stay in the cities where they first arrived — they spread! Often that spread is facilitated by our movement of firewood, plants, or outdoor household goods such as patio furniture.
The beech trees so important to wildlife conservation in the Northeast are under attack by two pathogens and at risk to an insect. Most alarming is the spread – in a dozen years! — of beech leaf disease DMF from Ohio to Maine. A leaf-feeding weevil is spreading south in eastern Canada. Please suggest that your Member or Senators to ask APHIS what steps it is taking to prevent the weevil’s introduction to the U.S.
‘Ōhi‘a trees make up 80% of the biomass of forests in both wet and dry areas of the Hawaiian archipelago. It is under attack by two diseases caused by introduced pathogens first detected in 2010. ‘Ōhi‘a forests support more threatened and endangered species than any other forest system in the U.S. They also play a uniquely important role in providing other ecosystem services, including water supplies.
Asking for the Money Pest Problems Deserve
To respond effectively to these pests and to the others that will be introduced in coming years, the key APHIS programs identified above must have adequate funds. The funding levels I request – and hope you will support – are lower than I would wish, but everyone expects the Congress to refuse significant increases in funding (see table at beginning of this blog).
The Tree and Wood Pests account supports eradication and control efforts targeting principally the ALB and spongy (= gypsy) moth. Eradicating the ALB normally receives about two-thirds of the funds. The programs in Massachusetts, New York, Ohio, and South Carolina must continue until eradication succeeds.
Oregon detected the EAB in 2022. Although the state and Portland have been preparing for a decade for this eventuality, there will still be significant impacts. Four percent of Portland’s street trees are ash – more than 9,000 trees. Young ash constitute three percent of young trees in parks. Loss of Oregon’s ash will also have severe ecosystem impacts. In Willamette Valley wetlands, ash constitutes up to 100% of the forest trees. Washington and California are also concerned. Indeed, the Hudgins study identified Seattle and Takoma as likely to lose thousands of ash trees. The numerous ash in riparian forests, windbreaks, and towns of North Dakota are also at risk since the EAB is established in South Dakota, Minnesota, and Manitoba.
APHIS manages damaging pests introduced on imported plants or other items through its Specialty Crops program. The principal example is its efforts to prevent spread of the SOD pathogen through the interstate trade in nursery plants. We noted above that this program is not as successful as it should be. We support the Administration’s request for $222 million; however, you might suggest that your Member or Senator urge APHIS to allot adequate funding under this budget line to management of SOD, rapid ʻōhiʻa death pathogens in Hawai`i, and beech leaf disease and elm zig-zag sawfly in the East.
The Pest Detection program is key to the prompt detection of newly introduced pests that is critical to successful pest eradication or containment. The “Methods Development” program enables APHIS to improve development of essential detection and eradication tools.
The Administration’s request include a $1 million emergency fund. This is far below the level needed to respond when a new pest is discovered. Funding constraints have hampered APHIS’ response to past pest incursions.
Please note that many of the members of the Agriculture Appropriations Subcommittee are from states where non-native pests are probably not top of mind. It is important that everyone that knows about these threats communicate with your Member/Senators!!
Members of House or Senate Subcommittees that Fund APHIS
(Names of Senators are italicized)
STATE
MEMBER
APHIS APPROP
HOUSE
SENATE
AK
Lisa Murkowski
X
AL
Jerry Carl Katie Britt
X
X
X
Calif
Barbara Lee David Valadao Josh Harder Diane Feinstein
X X X
X X X
X
FL
Debbie Wasserman Scultz Scott Franklin
X X
X X
GA
Sanford Bishop
X
X
ID
Mike Simpson
X
IL
Lauren Underwood
X
X
KS
Jerry Moran
X
X
KY
Mitch McConnell
X
X
LA
Julia Letlow Ashley Hinson
X X
X X
MD
Andy Harris Chris Van Hollen
X
X
X
ME
Chellie Pingree Susan Collins
X X
X
X
MI
John Moolenaar Gary Peters
X X
x
X
MN
Betty McCollum
X
X
MS
Cindy Hyde-Smith
X
X
MT
Jon Tester Ryan Zinke
X
X
X
NB
Deb Fischer
X
ND
John Hoeven
X
X
NM
Martin Heinrich
X
X
NV
Mark Amodei
X
OH
Marcy Kaptur
X
X
OR
Jeff Merkley
X
X
X
PA
Guy Reschenthaler
X
X
RI
Jack Reed
X
TX
Michael Cloud Jake Ellzey
X
X X
UT
Chris Stewart
X
VA
Ben Cline
X
X
WA
Dan Newhouse Derek Kilmer
X
X X
WV
Shelly Moore CapitoJoe Manchin
X
X X
WI
Mark Pocan Tammy Baldwin
X X
X
X
SOURCES
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. PNAS August 27, 2019. Vol. 116 No. 35 17371–17376
Haack R.A., J.A. Hardin, B.P. Caton and T.R. Petrice .2022. Wood borer detection rates on wood packaging materials entering the United States during different phases of ISPM#15 implementation and regulatory changes. Front. For. Glob. Change 5:1069117. doi: 10.3389/ffgc.2022.1069117
Hudgins, E.J., F.H. Koch, M.J. Ambrose, and B. Leung. 2022. Hotspots of pest-induced US urban tree death, 2020–2050. Journal of Applied Ecology
Leung, B., M.R. Springborn, J.A. Turner, and E.G. Brockerhoff. 2014. Pathway-level risk analysis: the net present value of an invasive species policy in the US. Front Ecol Environ 2014; doi:10.1890/130311
Liebhold, A.M., E.G. Brockerhoff, L.J. Garrett, J.L. Parke, and K.O. Britton. 2012. Live Plant Imports: the Major Pathway for Forest Insect and Pathogen Invasions of the US. Frontiers in Ecology.
MacLachlan, M.J., A. M. Liebhold, T. Yamanaka, M. R. Springborn. 2022. Hidden patterns of insect establishment risk revealed from two centuries of alien species discoveries. Sci. Adv. 7, eabj1012 (2021).
Szakonyi, M. 2023. Sourcing shift from China pulls US import share to more than a decade low.
The characteristic zigzag pattern Picture: Kelly Oten, NC State University.
Guest blog by Kelly Oten, NC State University
The elm zigzag sawfly [EZS; Aproceros leucopoda Takeuchi (Hymenoptera: Argidae)] is the newest invasive forest insect detected in the eastern US. The colloquially-used common name, currently going through the ESA common name approval process, is not only catchy, but perfectly describes this defoliator’s unique feeding damage. As EZS feeds on elm leaves, it weaves a zigzag pattern from the margin of the leaf towards the mid-vein.
An Expansive — and Quickly Growing – Range
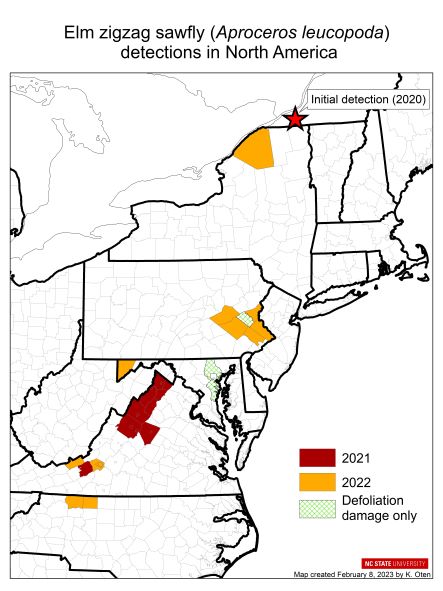
Native to East Asia, the first confirmation of EZS in North America occurred in August 2020 in Québec, Canada when an iNaturalist user posted a photo showing the characteristic zigzag defoliation. The observer realized it was potentially EZS and emailed local entomologists in the province who visited the site, collected specimens, and obtained species confirmation through the Canadian Food Inspection Agency Entomology Lab (Martel et al. 2021). However, this detection was not actually the beginning.
Three months before the Canadian detection, the same defoliation pattern was observed in Frederick County, Virginia, USA. Observers suspected EZS, but no specimens were recovered and therefore identification could not be confirmed. A year later, the site was revisited and this time, bingo—specimens were present and confirmed as EZS. Subsequent surveys that summer led to detections in eight additional Virginia counties. At the same time, the telltale defoliation was observed in Lehigh County, Pennsylvania, USA, but no specimen could be recovered for confirmation. In 2022, EZS popped up more widely; four additional states confirmed EZS: Pennsylvania, North Carolina, Maryland, and New York.
map created by Kelly Oten, NCSU
Though new to North America, this insect has a history of invasiveness. First detected in Europe in 2003, it now occupies areas from the United Kingdom and France in the west, to Russia and Kazakhstan in the east (Ashikbayev et al. 2018, iNaturalist 2022).
The strange and unusual biology of elm zigzag sawfly
Like all Hymenopterans, EZS goes through four life stages: egg, larva, pupa, adult. Eggs are laid along leaf margins; after hatching, larvae feed on leaf foliage in a zigzag pattern towards the mid-vein. Older larvae consume the leaf more entirely, leaving behind the mid-vein and thicker lateral veins only. Before pupating, larvae spin a cocoon within which they pupate. Cocoons are seasonally dimorphic; summer pupae (which emerge as adults in 4-7 days) are net-like and attached to leaves or twigs. Overwintering pupae are solid-walled and found in leaf litter or soil. Interestingly, overwintering pupae are not just produced from the last generation of the year. Even early in the summer, overwintering pupae may develop alongside summer pupae. Adults are able to begin oviposition immediately; not only do they not need to feed, but they don’t need to find a mate either! EZS is parthenogenetic, meaning they reproduce without mating. In fact, no male EZS has ever been recorded and it’s believed the species is entirely female.
As elm zigzag sawfly larvae (bottom left on leaf) grow, they feed more wholly on elm leaves. Picture: Kelly Oten, NCSU
The entirety of this life cycle can last ~20-36 days when not overwintering. However, the voltinism of this pest is highly variable. Papp et al. (2018) recorded up to seven generations in a year on lab-reared colonies, but in nature in Europe, anywhere between two and six generations has been recorded (Blank et al. 2010, Mol and Vonk 2015). In Virginia, two generations were recorded in 2021 and 1 in 2022. It is unknown what factors play into the number of generations per year, but it’s clear that it’s highly variable. The ability of EZS to multiply rapidly and have multiple generations per year suggest large populations can build in a relatively short time. In fact, this was observed in North Carolina in 2022 and in Europe several times before. Large populations are capable of severe defoliation and may cause long-term impacts on tree health.
This collective life cycle description is based on Blank et al. 2010, Martel et al. 2021, Martynov and Nikulina 2017, and Wu 2006.
Spread
EZS has astonished many with how it seems to be popping up all over the place in such a short amount of time. Since 2020, it has been detected in five US states and at several sites along the St. Laurence River in the Canadian province. The adults are strong fliers, capable of spreading 45-90 km (~27-55 mi) per year (Blank et al. 2014). Given the fact they’re parthenogenetic, relatively small numbers can disperse to begin new populations. Of perhaps greater concern is the potential for long-range dispersal. In heavy infestations in North Carolina, cocoons were found not just attached to leaves and stems, but also non-living objects, suggesting a possible mechanism of long-range dispersal should they become attached to vehicles or other objects transported long distances. In addition, EZS damage ranges from minor to severe defoliation. When populations are low and feeding is minor, it’s less likely to be detected unless intentional surveys are conducted. This cryptic nature might suggest it’s in more places than we are currently aware.
An elm zigzag sawfly cocoon attached to a fence post. Picture: Kelly Oten, NCSU
So, what’s the big deal?
In short, we don’t know yet. Generally speaking, defoliation by insects causes little long-term harm to tree health but severe and/or repeated defoliation can weaken or sometimes kill a host. In Europe, trees severely defoliated by EZS are typically able to re-leaf but may suffer branch dieback and/or reduced growth (Blank et al. 2010, Zandigiacomo et al. 2011). Also of note, EZS is attacking elm, an already-threatened tree due to widespread mortality cause by Dutch elm disease in the 1900s. Defoliation by EZS could further weaken infected trees or, at the very least, present an additional threat for remaining elms (Blank et al. 2010). While it seems aesthetic damage will be the primary concern with EZS, the potential for long-term tree health impactsin the US is uncertain and should be investigated. For now, anyone that finds EZS or its characteristic defoliation pattern are encouraged to report it to their respective state agriculture or forestry agency.
A row of winged elm (Ulmus alata) in NC were severely defoliated by elm zigzag sawfly. Picture: Kelly Oten, NCSU
[See Faith’s earlier blog about the zigzag sawfly here.]
References
Ashikbayev, N. Z., N. S. Mukhamadiyev, G. Z. Mengdibayeva, M. B. Temirzhanov, and N. K. Kuanyshbaev. 2018. Development of forest entomology in Kazakhstan, pp. 42–47. In T. I. Espolov, K. M. Tireuov, E. I. Islamov, S. B. Baizakov, K. T. Abayeva, E. Z. Kentbaev, and B. A. Kentbaeva (eds.), Actual problems of sustainable development in forestry complex, vol. 2. Aitumar Publishing, Almaty, Kazakhstan.
Blank, S. M., H. Hara, J. Mikulás, G. Csóka, C. Ciornei, R. Constantineanu, I. Constantineanu, L. Roller, E. Altenhofer, T. Huflejt, and G. Vétek. 2010. Aprocerosleucopoda (Hymenoptera, Argidae): an East Asian pest of elms (Ulmus spp.) invading Europe. Eur. J. Entomol. 107: 357–367.
Blank, S. M., T. Köhler, T. Pfannenstill, N. Neuenfeldt, B. Zimmer, E. Jansen, A. Taeger, A.D. Liston. 2014. Zig-zagging across Central Europe: recent range extension, dispersal seed and larval hosts of Aprocerosleucopoda (Hymenoptera, Argidae) in Germany. J. Hymenopt. Res. 41: 57-74.
iNaturalist. Available from https://www.inaturalist.org. Accessed August 2022.
Martel, V., O. Morin, S. Monckton, C. Eiseman, C. Béliveau, M. Cusson, and S. Blank. 2021. Elm zigzag sawfly, Aproceros leucopoda (Hymenoptera: Argidae), recorded for the first time in North America through community science. Can. Entomol. 154: E1.
Martynoz, V. V., and T. V. Nikulina. 2017. Population surge of zigzag elm sawfly (Aproceros leucopoda (Takeuchi, 1939): Hymenoptera: Argidae) in the Northern Ciz-Azov Region. Russ. J. Biol. Invasions 8: 25-34.
Mol, A. W. M., and D. H. Vonk. 2015. De iepenzigzagbladwesp Aproceros leucopoda (Hymenoptera: Argidae), een invasieve exoot in Nederland. Entomol. Ber. 75: 50-63.
Papp, V., M. Ladányi, and G. Vétek. 2018. Temperature-dependent development of Aproceros leucopoda (Hymenoptera: Argidae), an invasive pest of elms in Europe. J. Appl. Entomol. 142: 589-597.
Wu, X. Y. 2006. Studies on the biology and control of Aproceros leucopoda. Plant Prot. 32: 98-100.
Zandigiacomo, P., E. Cargnus, and A. Villani. 2011. First record of the invasive sawfly Aproceros leucopoda infesting elms in Italy. Bull. Insectology 64: 145-149.
nearly dead ash in Shenandoah National Park; photo by F.T. Campbell
I participated in the annual USDA Interagency Invasive Species Research Forum in Annapolis in January 2023; as usual, I learned interesting developments. I focus here on updates re: efforts to protect ash and hemlock
Hopeful Developments re: countering EAB to protect ash
There are hopeful results in both the biocontrol and resistance breeding programs. The overall goal is to maintain ash as a viable part of the North American landscape.
Biocontrol
Juli Gould (APHIS) reminded us that the agency began a classical biocontrol program targetting emerald ash borer (EAB) in 2003 – only a year after EAB had been detected and much earlier than is the usual practice. [Thank you, former APHIS PPQ Deputy Administrator Ric Dunkle!] By 2007 scientists had identified, tested, and approved three agents; a fourth was approved in 2015.
Nicole Quinn (University of Florida) stressed that the egg prarasitoid, Oobius — if it is effective — could prevent EAB from damaging trees. However, it is so small that it is very difficult to sample. One small study demonstrated that Oobius will parasitize EAB eggs laid in white fringe trees (Chionanthus virginicus) as well as in ash. This is important because it means this secondary host is not likely to be a reservoir of EAB.
The numbers
According to Ben Slager (APHIS), more than 8 million parasitoids have been released at 950 sites since the program began in 2007. These releases have been in 418 counties in 31 states, DC, and four Canadian provinces. Still, these represent just 28% of infested counties. Parasitoids have been recovered in 21 states and two provinces.
Rafael de Andrade (University of Maryland) specified that these releases included more than 5 million Tetrastichus in 787 sites; ~2.5 million Oobius in 828 sites in 30 states; ~500,000 Spathius agrili – lately only north of the 40th parallel. Releases of Spathiusgalinae began in 2015; so far ~ 470,000 in 395 sites.
Impact
Several presenters addressed questions of whether the agents are establishing, dispersing, and – most important – improving ash survival. Also, can classical biocontrol be integrated with other management techniques, especially use of the pesticide emamectin benzoate.
Dispersal
Several studies have shown that the four biocontrol agents disperse well (with the caveat that Oobius is very difficult to detect so its status is much less certain).
Implementation considerations
De Andrade found that the longer the delay between the date when EAB was detected and release of Oobius, the less likely Oobius will be recovered. Tetrastichus surprised because the higher the numbers released, the fewer were recovered. He could determine no association between recovery of S. agrili and variations in release regime [numbers released; delay in releasing biocontrol agents; or frequency of releases]. He said it is too early to assess Sp. galinae since releases began only in 2015, but he did see expected relationship to propagule pressure – the more wasps released, the higher the number that were recovered. Sp. galinae did surprise in one way: it seemed to perform better at lower latitudes. De Andrade noted he was working data from less than half of release sites. He asked collaborators to submit data!!!!
Initial signs of ash persistence and recovery
Claire Rutledge (Connecticut Agriculture Experiment Station) determined that
More large trees were surviving in plots where the biocontrol agents were released
EAB density was lower at long-invaded sites
Parasitism rates were similar across release age treatments and release/control plots
Gould focused on protecting saplings so they can grow into mature trees which could be sources of seeds to establish future generations. She noted that there are many “aftermath” forests across the northern United States – those dominated by ash saplings.
In Michigan, at a site of green ash, as of 2015 – 2021, EAB populations are still low, parasitism rate by Tetrastichus and S. galinae high. The percentage of saplings that remained healthy was greater than 80%. There were similar findings in white ash in New York: very low EAB larval density; and more than 70% of ash saplings had no fresh galleries. Gould reported that Tetrastrichus impcts could be detected within three years of release.
So, EAB are being killed by the biocontrol agents combined with woodpecker predation; but in their fourth instar, after considerable damage to the trees.
downy woodpecker in Central Park, NYC. photo by Steven Bellovin, Columbia University
Jian Duan reported on two long-term studies in green & white ash in Michigan and New England. His team used the most labor-intensive but best approach to determine EAB larval mortality and the cause – debarking trees – to determine whether the EAB larva were parasitized, were preyed on by woodpeckers, or were killed by undetermined cause, such as tree resistance, disease, or competition. In Michigan, he linked a crash of EAB population in 2010 was caused by Tetrastichus; EAB tried to recover, but crashed again, due to S. galinae. EAB larval densities had been reduced to 10 / m2. Predation by abundant woodpeckers and the native parasitoid Atanycolus was also important.
In New England, EAB has also declined from 20-30 larvae /m2 to ~ 10 m2.
In Michigan, healthy ash with dbh of larger than 5 inches were much more plentiful in sites where parasitoids had been released. Their survival/healthy rate also was much higher in release sites but the difference declined as years passed. In New England there were growing numbers of healthy trees in 2021-22; (almost none in 2017). Duan conceded that he could not prove a direct link but the data points to recovery.
Tim Morris (SUNY-Syracuse) found that white ash saplings continued to die in large numbers, but the mortality rate was significantly below the rate in 2017. Canopy conditions varied; some trees that were declining in 2013 were recovering in 2017. Forty percent of “healthy” ash in 2013 continued recovering in 2021. Few living trees were declining; trees were either healthy or dead. He thinks probably a combination of genetics and presence of parasitoids explains which trees recover. Morris also reported some signs of regeneration.
beaver feeding on ash saplings, Fairfax County, Va; photo by F.T. Campbell
At this point, I noted that in parts of northern Virginia, beavers have killed ash saplings. Morris reported finding the same in some sites in New York. Perhaps others have, also; my comment was greeted by laughter.
Theresa Murphy (APHIS) looked at integration of biocontrol and insecticide treatment in urban and natural sites. A study of black and green ash in Syracuse, NY Naperville, IL, and Boulder, CO found continued high parasitism by Tetrasticus and S. galinae and woodpecker attacks in trees treated with emamectin benzoate. Researchers could not detect Oobius. By 2020, most of the untreated trees had died but treated trees remained healthy.
Murphy has begun studying integration of biocontrol and pesticides in green and black ash forests. The goal is to protect large trees to ensure reproduction; the biocontrol agents do not yet protect the large trees. This is especially important for black ash because it declines very quickly after EAB invades. Sites have been established in New York, through collaboration with New York parks, Department of Environmental Conservation, and the Mohawk tribe. She is still looking for sites in Wisconsin – where EAB is spreading more slowly than expected.
1 of the infested ash in Oregon; photo by Wyatt Williams, ODF
Max Ragozzino of the Oregon Department of Agriculture reported on imminent release of biocontrol agents targetting the recently detected outbreak there. I am encouraged by the rapid response by both the state and APHIS.
EAB resistance in ash
Jennifer Koch (USFS) said the goal is not to produce populations where every seedling is fully EAB-resistant, but to develop populations of ash trees with enough resistance to allow continued improvement through natural selection while retaining sufficient genetic diversity to adapt to future stressors (changing climate, pests, diseases). The program has developed methods to quantify resistance in individuals.. Initial field selections of “lingering ash” were shown to be able to kill as many as 45 % of EAB larvae. Already green ash seedling families have been produced by breeding lingering ash parents. This first generation of progeny had higher levels of resistance, on average, than the parent trees. Each generation of breeding can increase the proportion of resistance. Although the bioassays to test for EAB-resistance are destructive (e.g., cutting and peeling to count numbers of surviving larvae), the potted ash seedling stumps can resprout. Once the new sprouts are big enough they are planted in field trials to correlate bioassay results with field performers. Poor performers are culled; those with higher levels of resistance remain and become sources of improved seed.
To ensure preservation of local adaptive traits, this process must be repeated with new genotypes to develop many seed orchards from across the species’ wide range. To support this work, concerned scientists are building multi-partner collaborative breeding networks. These organizations provide ways for citizens and a variety of partners to engage through monitoring and reporting lingering ash, making land available for test planting, and helping with the work of propagation.
Resistance levels in some of the first generation progeny were high enough for use in horticulture, where it is important that trees can remain healthy in challenging environments (street trees, city parks, landscaping, etc.). Koch hopes to develop about a dozen cultivars comprising the best-performing trees, appropriate for planting in parts of Ohio, Michigan, Indiana, and Pennsylvania. Local NGO partners are planting some of these promising genotypes in Detroit to see how they withstand EAB attack.
a black ash swamp; photo via Flickr
The threat to black ash is especially severe, and this species presents unique difficulties. While scientists found several seedlings from unselected seedlots had killed high levels of larvae, those deaths did not always result in better tree survival. Koch thinks the tree’s defense response becomes detrimental to tree by blocking transport of water and nutrients. She is working with experts in genomics and others, such as Kew Royal Botanic Gardens, to try to identify candidate trees for breeding programs. The genomics work has been supported by APHIS and the UK forest research agency, DEFRA. Michigan and Pennsylvania have supported the breeding work. USFS Forest Health Protection has supported work with black and Oregon ash (see below) (J. Koch, USFS, pers. comm.).
Koch has also begun working with Oregon ash, in collaboration with the USFS Dorena Genetic Resource Center (located in Cottage Grove, Oregon) and other partners.
dead hemlock in Massachusetts; photo by Ian Kinahan, University of Rhode Island
Scientists are still trying to find the right combination of biocontrol, chemical treatments, and silvicultural manipulation.
For several years, hope has focused on two has been on two predatory beetles, Laricobius nigrinus and L. osakiensis. Scott Salom (Virginia Tech) reports that release of these beetles over the past 20 years has had a significant impact on HWA density and tree photosynthetic rate and growth. However, Laricobius aredifficult to rear and they attack only the sistens generation of the adelgid. Ryan Crandall (University of Massachusetts) reports it has been difficult to establish these beetles in the Northeast. He links this difficulty is caused by temporary drops in HWA populations after cold snaps.
Scientists now agree that need to find predators that attack HWA during other parts of its lifecycle. Hope now focuses on silverflies — Leucotaraxis argenticollis and Le. piniperda. While both species are established in eastern North America, the clades in the east feed almost exclusively on pine bark adelgid, and have not begun attacking HWA. Biocontrol practitioners therefore collect flies in the Pacific Northwest for release in the east. Salom is increasing his lab’s capacity to rear silverflies and exploring release strategies.
Preliminary evidence indicates that the western clades of Leucotaraxis are establishing, although data are not yet definitive (Havill, USFS).
Detecting the presence of biocontrol agents presents several challenges. Tonya Bittner (Cornell) described efforts to use eDNA analysis for this. Some puzzles have persisted; e.g., at some sites, she detected eDNA but caught no silverflies. This raised the question of long eDNA associated with the original release might persist. Another problem is that the assay cannot separate the introduced western L. nigrinus from the native congener, L.rubus (which also does not feed on HWA). She continues efforts to improve this technique.
Others explored interactions of the biocontrol agents with insecticides. Salom is studying the impact of soil-applied insecticides on Laricobius populations, which aestivate in the soil. Preliminary results showed significant reduction in the beetle’s population under soil drench application but not under soil injection. He has not yet analyzed all the data.
Michigan is trying to prevent spread of HWA from five counties along the eastern shore of Lake Michigan (where HWA was introduced on nursery stock) to widespread hemlock forests in northern part of the state. Phil Lewis (APHIS) is studying persistence of systemic insecticides in hemlock tissues, particularly twigs and needles. The pesticides involved are imidacloprid, dinotefuran, and Olefin. He has found that pesticide levels are highest 18 – 22 months after treatment, then decline. They are significantly higher after trunk injection compared to bark spray or soil treatments. Imidacloprid had higher residues in twigs; dinotefuran in needles. This difference affects the likelihood of adelgids actually ingesting the toxin.
healthy hemlock in experimental gap; Jefferson National Forest, VA; photo by Bud Mayfield, USFS
Bud Mayfield (USFS) reported on his study of silvicultural strategies to support healthier hemlocks. While hemlocks normally thrive in shade, it has been determined that sunlight assists small trees reducing HWA sufficiently to counter the tree’s leaf-level stress. Small sapling hemlocks grown in sunlight fix more carbon and convert it to growth in shoots and trunk diameter.
Mayfield found promising immediate suppression of HWA in large gaps in Georgia and Tennessee. By the third year the saplings were still growing, although their faster growth had attracted more HWA. These findings were less clear farther north in central Virginia and western Maryland – Mayfield thinks because HWA pressure there is lower. However, managers must maintain the gaps by cutting rapidly-growing competing woody species. He plans to test this strategy farther north in Pennsylvania. He is still trying to determine the optimal size of the gap.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
The Old Man – a giant ash tree in Wytham Woods; photo from https://theoldmanofwytham.com/2018/11/29/ash-dieback-in-wytham-woods/
I campaign for protecting trees – especially trees growing to their natural capacity in the habitats in which they have evolved. I focus on the threat to these trees from non-native insects and various pathogens (fungi, nematodes …). I have often expressed my distress because others appear to place a low priority on this goal. I have also asked whether protecting trees might be given a higher priority by more decision-makers if they recognize trees’ vitally important role in countering climate change.
For this reason, I have blogged several times about studies examining the role trees play in sequestering carbon — see here & here & here.
A new study demonstrates that protecting large, old trees – almost by definition in their natural environment – is vitally important. Planting new, small, trees is helpful but cannot substitute for the venerable trees.
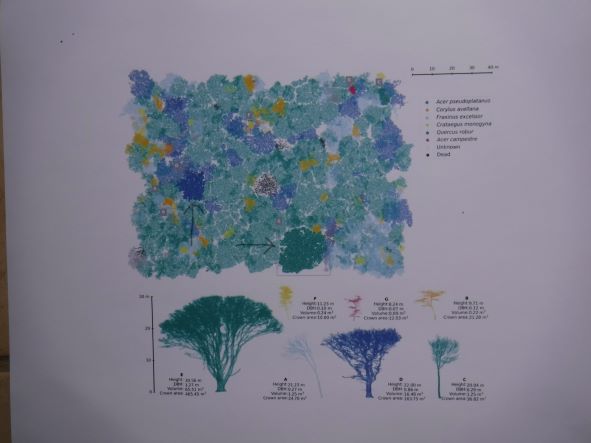
Calders and colleagues (full citation at the end of the blog; open access!) have used new technology to update assessments of the amount of carbon sequestered in trees. They conducted their study in a temperate hardwood forest – Wytham Woods, a typical broadleaf temperate forest in Oxfordshire, southern Great Britain. [Wytham Woods is also the site of two of the “Inspector Morse” mysteries – “Secret of Bay 5B” and “A Way Through the Woods”.]
They found that these trees sequester 1.77 times more carbon in their above-ground biomass (AGB) than previously believed based on currently-used models.
One consequence of their findings is that countries using the standard assessment method (which was developed by Robert Bunce in 1968) are reporting inaccurate carbon sequestration estimates to the United Nations per the Paris climate accords. (Calders et al. believe that calculations for conifer species are probably more accurate than those for deciduous forests.)
A second consequence is that death of large trees – from whatever cause – will result in greater loss of carbon storage than previously thought.
Old v. New Measurements
The underlying Bunce dataset and algorithm applied in most European biomass estimates were based on a small sample: 200 trees belonging to five taxa growing in one forest area. The models were derived by cutting down trees and weighing them to determine tree biomass. Smaller trees were used because they are easier to process. The scientists then extrapolated the biomass of bigger trees based on the assumption that correlation between tree size and mass is independent of tree size. This assumption has rarely been tested because of the difficulty and expense of carrying out this type of destructive sampling.
The higher estimates of carbon storage in Calders et al. arise in part from the bias towards small trees in calibration of the earlier models. Calders et al. found that trees do not follow a size-invariant scaling relationship, particularly at larger size; it is important to include crown area. Thus, Calders and colleagues calculated a higher sequestration rate for trees in Wytham Woods that fell within the size range used in developing the Bunce allometric model.
In addition, changes in forest management have increased the abundance of larger trees compared to the 1960s when Bunce carried out his study. Indeed, many of the trees in Wytham Woods are nearly twice as large as the trees used in the original calculation of biomass. The median dbh in Bunce (1968) is 8.4 cm; the mean dbh for the TLS dataset (based on a 2015 inventory) is 15.9 cm. The large trees represent a high proportion of the above-ground biomass: 50% of AGB in Wytham Woods was associated with fewer than 7% of the trees (those with dbh greater than 53.1 cm). All these trees were larger than the trees used to calibrate the widely used allometric model.
Calders et al. say that the distribution of tree size (trunk diameter) in Wytham Woods is representative of broadleaved species throughout Great Britain. Basal area had doubled in 40 years from 1974. Thus, the growth trajectory reflected at Wytham Woods – and presumably across Britain – resulted in a net carbon sink of ~1.77tha-1year-1ha in Calder et al’s 3D analysis. This is almost double the ~1tha-1year-1ha derived using the traditional allometric models. .
Methodology
graphic from Calders et al. large maple (green) and oak (blue) trees illustrated by LiDAR images – profiles and location in the forest (indicated by arrows); copyright Ecological Solutions and Evidence
Calder et al. used terrestrial laser scanning (TLS; terrestrial LiDAR) methods & 3-dimensional analysis to derive tree volume and convert this to above-ground biomass (AGB) and carbon sequestration. They scanned 815 live standing trees in Wytham Woods during winter so leaves did not complicate computations. They found:
total volume of these 815 trees was 742.6±3.9m3ha-1.
TLS-derived AGB = 409.9tha-1. This is significantly greater than the 231.9tha-1 resulting from applying the Bunce allometric models.
In sum, 1.77 times more carbon is stored per ha according to this model than carbon values derived through the allometric AGB models developed by Bunce.
Calder et al. describe the threat to European carbon sequestration projections caused by ash dieback. Ash dieback has been spreading across Europe since the 1990s – although the causal agent was not determined until 2006 (Paap et al.). It is killing European ash across the continent. Some of these trees are large – that is, store impressive amounts of carbon. In Wytham Woods specifically, ash dieback threatens some of the largest trees.
Ash dieback disease was first observed in the United Kingdom in 2012; it reached Wytham Woods in 2017. Ash contributed ~13.2% of the biomass carbon sequestration in the study area. However, the species’ presence in all of Wytham Woods might approach ~34%. Ash comprised 75% of seedlings in 2012. Ash is one of three species that contribute >26% of broadleaved tree AGB & carbon for Great Britain as a whole. The British Woodland Trust expects the UK to lose 80% of its ash trees. As a result, Wytham Woods, Britain, and, by extension, a significant amount of European temperate deciduous forests, are likely to become a substantial carbon source in the next decades.
A dead elm tree on Skelston Moor; photo by Walter Baxter; CC BY-SA 2.0
I note that Europe has already lost any sequestration benefits it would have enjoyed from large elm trees due to “Dutch” elm disease. Various Phytophtoras are killing trees in Britain and Ireland.
I recently described threats to plane trees, pines, and other trees across Europe.
I interpret these findings as demonstrating that protecting large trees growing in natural ecosystems is highly important as we try to cope with climate change. This will require determined, sustained, and strategic actions in the face of disturbances predicted to increase as result of changes in climate and the human activities that contribute to climate change – e.g., overexploitation of natural resources, conversion of natural systems to human use, shipping goods around the globe, …
Calders and colleagues say we cannot afford to lose substantial reservoirs of carbon currently sequestered in temperate forests. Such forests currently account for ~14% of global forest carbon stocks in their biomass and soil. Their importance is growing because of widespread deforestation in the tropics.
What is To Be Done? (to cite Lenin)
Calders and colleagues call for several actions to address potential biases in biomass carbon estimates and drastically improve estimates of forest biomass:
(i) Research to improve knowledge about carbon sequestration levels in trees. This will require
a) greater sampling using such nondestructive methods as TLS to estimate AGB of a wider variety of forest types,
b) improved understanding of wood density, and
c) properly testing the fundamental assumption of size dependency in allometric models.
(ii) Develop empirical models of AGB that do not assume size invariance. This might require. This implies more destructive harvesting to obtain data from a variety of forest compositions, locations, etc,
(iii) Establish a biomass reference network of permanent sample plots specifically designed for estimating AGB. The improved data can then be fed into satellite-derived biomass estimates, which are likely to become the de facto standard for assessing the state and change of forest AGB at large scales. The GEO-TREES database can help. It aims to build on existing long-term ecological plot networks, by including TLS, airborne laser scanning & other ancillary data (including harvest measurements) to specifically allow for upscaling of AGB & development of new empirical models.
(iv) Ensure much better traceability in the use of allometric models. If applying a model to a site at several removes from the original data, e.g., published allometric models, clearly identify where and when the underpinning data were collected, the number and size range of trees from which models were derived, and clarify any assumptions regarding environmental conditions, wood density etc. Database initiatives such as GlobAllomeTree can help.
North American Situation
remains of Michigan’s champion green ash
A study in 2019 (Fei et al. 2019; full citation at end of the blog) has already estimated that 41% of total live (woody) biomass in forests of the “lower 48” states is at risk from the most damaging of introduced pests. The greatest biomass loss was caused by emerald ash borer, Dutch elm disease, beech bark disease, and hemlock woolly adelgid. Before arrival of these non-native pests, mature ash, elms, beech and hemlock were large – providing significant storage of carbon (and other ecosystem services). A complication is that elms and beech, at least, began dying decades before the underlying (Forest Inventory and Analysis; FIA) data began to be collected. Consequently, the reported mortality rates underestimate the actual loss in biomass associated with these pests.
Did Fei et al. rely on biomass estimates based on measurements and algorithms now questioned by Calders et al.? One of the co-authors, Dr. Randall Morin, has told me that USFS scientists are shifting to new models that will result in a slight bump in overall biomass for the U.S. largely because of increased recognition of the biomass in crowns and limbs. However, the new models are based partly on a felled-tree study, so I wonder if they will have similar issues.
Certainly in some situations that threat posed by non-native pests is not yet being adequately incorporated. Badgley et al. (2022) analyzed the California cap-and-trade program to determine whether forest projects enrolled under its provisions can provide sufficiently permanent carbon sequestration. They determined that sequestration losses tied to mortality of one tree species (tanoak; Notholithocarpus densiflorus) due to one disease – sudden oak death – would fully deplete the “buffer pool” set aside to compensate for losses due to disease and insect infestations. This leaves the program unable to provide the promised benefits in carbon sequestration. SOD continues to spread and tanoaks (and other tree species) to die. California along is home to other tree-killing pathogens and insects, e.g., white pine blister rust, Port-Orford cedar root disease, Fusarium dieback, goldspotted oak borer …
California live oak killed by GSOB; photo by F.T. Campbell
Furthermore, the program allows enrollment of forests across the United States, so the multiple pests threatening ash, hemlocks, oaks, and other tree taxa across North America must also be accommodated. I have not even mentioned the likelihood that additional tree-killing pests will be introduced in the future.
How can scientists enhance the credibility of well-intentioned efforts to incorporate forest conservation into strategies aimed at mitigating climate change?
[A separate study by Oxford University has estimated that 2 billion tonnes of CO2 are removed from the atmosphere every year – 99% of it by trees. They point out that this is not sufficient to help Earth avoid temperatures rising above Paris-set levels. See an article by Lottie Limb, Reuters, published 19 January 2023 (sorry – I don’t have a direct link).]
SOURCES
Badgley, G., Chay, F., Chegwidden, O.S., Hamman, J.J., Freeman J. and Cullenward, D. 2022. Calif’s forest carbon offsets buffer pool is severely undercapitalized. Front. For. Glob. Change 5:930426. doi: 10.3389/ffgc.2022.930426
Bunce, R. G. H. (1968). Biomass and production of trees in a mixed deciduous woodland: I. Girth and height as parameters for the estimation of tree dry weight. Journal of Ecology, 56, 759–775.
Calders, K., H. Verbeeck, A. Burt, N. Origo, J. Nightingale, Y. Malhi, P. Wilkes, P. Raumonen, R.G.H. Bunce, M. Disney. Laser scanning reveals potential underestimation of biomass carbon in temperate forest. Ecol Solut Evid. 2022;3:e12197. wileyonlinelibrary.com/journal/eso3 open access!
Paap, T., M.J. Wingfield, T.I. Burgess, J.R.U. Wilson, D.M. Richardson, A. Santini. 2022. Invasion Frameworks: a Forest Pathogen Perspective. FOREST PATHOLOGY Current Forestry Reports https://doi.org/10.1007/s40725-021-00157-4
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Phytopthora ramorum-infected potted plants; photo by Washington State University
At this year’s USDA Invasive Species Forum I will be seeking to promote a discussion of what American and other stakeholders can do to suppress spread of forest pathogens. I have raised this issue many times before. To see my blogs about the P4P pathway, scroll down below the archives to the “categories”. See especially here and here.
I note that:
Non-native invasive pathogens and pests are decimating forests worldwide, threatening biodiversity & limiting efforts to rely on forests to alleviate impacts of climate change.
Many of the most damaging non-native organisms are pathogens that are especially difficult to detect at borders or to contain or eradicate once introduced.
A principal pathway by which pathogens are introduced is the international trade in living plants, or “plants for planting” (P4P).
Forest pathologists have long advocated a more pro-active approach – but national and international plant health officials have not taken up the challenge. [think Clive Brasier, Bitty Roy, Thomas Jung, Michael Winfield …]
Austropuccinia psidii on Melalecua in Australia; John Tann via Flickr
At the global level I suggest that we need:
National agricultural agencies, stakeholders, FAO & International Plant Protection Convention (IPPC) to consider amending IPPC requirement that scientists identify a disease’s causal agents before regulating it. I think experience shows that this policy virtually guarantees that pathogens will continue to enter, establish, & damage natural and agricultural environments.
National governments & FAO / IPPC to fund greatly expanded research to identify microbes resident in regions that are important sources of origin for traded plants, vulnerability of hosts in importing countries, and new technologies for detecting pathogens (e.g., molecular tools, volatile organic compounds [VOCs]).
Researchers & agencies to expand international “sentinel plants” networks; incorporate data from forestry plantations, urban plantings, etc. of non-native trees.
Application of ISPM#36 to promote use of HACCP programs for plants in trade. (See also my discussion in Fading Forests III – link at end of this blog.)
‘ohi‘a trees killed by rapid ‘ohi‘a death; photo by Richard Sniezko, USFS
We Americans need to
Evaluate efficacy of current regulations – incorporating NAPPRA & Q-37 revision. Rely on AQIM data. Include arthropods, fungal pathogens, oomycetes, bacteria, viruses, nematodes. Include threats to U.S. tropical islands (Hawai`i, Puerto Rico, Guam, etc.) which are centers of plant endemism.
Apply existing programs (e.g., NAPPRA, Clean Stock Network, post-entry quarantine) to strictly regulate trade in plant taxa most likely to transport pests that threaten our native plants; e.g., plants belonging to genera shared between North American trees & plants on other continents.
Recognize that plant nurseries are incubators for microbial growth, hybridization, and evolution; require nurseries to adopt sanitary operation procedures regardless of whether they sell in inter-state or intra-state commerce
I will explain my sense of urgency by noting the many recent introductions of pathogens – most probably via P4P or cut vegetation:
13 outbreaks of Phytophthora-caused disease in forests and natural ecosystems of Europe, Australia and the Americas. Three of four known strains of P. ramorum are established in U.S. forests.
Myrtle rust (Austropuccinia psidii) has been introduced to 27 countries, including the U.S., Australia, and South Africa.
Two new species of Ceratocystis (C. lukohia & C. huliohia)—causal agents of rapid ‘ohi‘a death (ROD) – spreading on the Hawaiian Islands. The former species appears to have originated in the Caribbean; the latter in Asia.
Since 2012, beech leaf disease has spread from northeastern Ohio to Maine.
Boxwood blight (caused by 2 ascomycete fungi, Calonectria pseudonaviculata & C. henricotiae) introduced to at least 24 countries in 3 geographic areas: Europe / western Asia; New Zealand, North America.
ash dieback fungus (Hymenoscyphus fraxineus) has spread across Europe after introduction from Asia.
What do you think? Can we find more effective methods to curtail introductions?
beech leaf disease
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Platanus orientalis in Turkey; photo by Zeynek Zebeci
A current issue of the journal Forests (2022 Vol. 13) is a special issue focused on forest pests. This topic was chosen because of increased pest incursions. Choi and Park (full citations at the end of the blog) link this to climate change and increased international trade, as well as difficulties of predicting which pests will cause damage where.
The journal issue contains 15 papers. Several patterns appear throughout. First is the important role of international trade in living plants – “plants for planting” – in introductions. This is hardly news! A second pattern is that at least two North American species were introduced to Europe during the 1940s, probably in wood packaging used to transport military supplies during World War II.
This compilation provides the opportunity to review which organisms of North American origin have become damaging invaders in Eurasia — and sometimes other continents. For example, the journal carries four articles discussing pine wilt disease (PWD). It is caused by the North American nematode Bursaphelenchus xylophilus, and is vectored by wood-boring insects in the genus Monochamus. Beetles introduced from North America and those native to the invaded area are both involved. This disease is considered a severe threat to forest health globally. No apparent association with WWII exists for PWD.
Two fungal pathogens from North America cause serious damage in urban and natural forests of Europe and central Asia. Neither is discussed in the special issue:
Ceratocystis platani has devastated urban trees in the Platanus genus, especially the “London plane” hybrid, and the native European tree, Platanus orientalis. This fungus was accidentally introduced to southern Europe during WWII – as were the two insects described by Musolin et al. It was first reported in northern Italy and Mediterranean France in the early 1970s, but disease symptoms had been observed years earlier. C. platani is established across the northern rim of the Mediterranean and to the east in Armenia and Iran. The worst damage has been in Greece, especially in natural forest stands in riparian areas. Spread of the pathogen there is facilitated by root grafts and by tree wounds caused by floating wooden debris during floods (Tsopelas et al. 2017.)
Platanus orientalis along Voidomatis River in Greece; photo by Onno Zweers, via Wikimedia
Heterobasidion irregulare infects conifers. It has spread and killed large numbers of Italian stone pine (Pinus pinea). The disease was inadvertently introduced to central Italy in the 1940s. H. irregulare has greater sporulation potential and decays wood more quickly than the native congener H. annosum. H. irregulare appears to be replacing the European species; scientists fear it will exacerbate tree infection and mortality rates (Garbelotto, Leone, and Martiniuc. date?)
A third North American pathogen, sooty bark disease (Cryptostroma corticale) has been introduced to Europe. This disease, found on sugar maple in eastern North America, was detected in Great Britain in 1945; it is now throughout Europe (Tanney 2022). EPPO reports that it is widespread in western Europe and in some Balkan countries. The website provides no information on its impact in Europe.
Pests in Russia
A paper authored by Musolin, et al. discusses 14 species of invasive or emerging tree pests found in Russian forest and urban ecosystems. Of these, two are native to North America. Another eight pose a threat to North America if they are introduced here.
As Musolin et al. point out, Russia covers a huge territory across Europe and Asia – stretching 10,500 km, or 6,500 miles. These encompass a great variety of ecological zones. Russia is also actively involved in international trade. It is not surprising, then, numerous non-native organisms have been introduced.
As of 2011, 192 species of phytophagous non-native insects from 48 families and eight orders were documented in the European part of Russia. This number does not include the vast areas in Asian Russia. Additional introductions have probably occurred in the most recent decade. Some of these introduced species have cause significant economic losses. Still, Russia appears to rarely mount a serious control effort.
Of course, the opposite is also true: pests native to some part of Russia can be transported to new regions of Russia or beyond its borders. We North Americans have focused on various species of tussock moths (Lymantria spp., etc.). There are many others. Musolin et al. describe eight in detail. All the information in this blog are from that article unless otherwise indicated.
Two North American Species’ Damage in Eurasia
Both these introductions were detected around the year 2000. Was there some event – other than simply expanding trade – that might explain these introductions?
Leptoglossus occidentalis; photo by nutmeg66 via Flickr
Western Coniferous Seed Bug, Leptoglossus occidentalis
This insect from western North America has invaded Eurasia, North Africa, and Central America. The first detection in Europe was in 1999 in Italy. It spread quickly and is present now from Morocco to Japan, as well as in South Africa and South America. The seed bug is spreading northward in European Russia, including into the forest-steppe zone. Its ability to spread to the East is uncertain.
L. occidentalis attacks a wide range of Pinaceae and Cupressaceae. In the Mediterranean region it has had serious impacts on the pine nut supply (Ana Farinha, IUFRO, Prague, September 2021). In southern parts of Russia it has caused “significant damage”. L. occidentalis also vectors a pathogenic fungus Sphaeropsis sapinea (=Diplodia pinea), which causes diplodia tip blight. The cumulative damage of insect and pathogen to pines can be significant.
The introduction pathway to Russia is unknown. It might have flown from established populations in Europe, or it might have been transported on plants for planting or Christmas decorations.
Oak Lace Bug, Corythucha arcuata
This insect is widespread in the United States and southern Canada. It was first detected in Europe – again, Italy – in 2000. Twenty years later it has spread to almost 20 countries.
Russia was invaded relatively recently; the first outbreak was detected in 2015 in the subtropical zone along the Black Sea coast and Caucasus. Musolin et al. expect the lace bug to spread to natural forests of Central Asia and other countries of the Caucasus. Its spread will be assisted by air currents and movement of plants for planting. The insect is causing considerable aesthetic damage, but other impacts have not been estimated.
Hosts include many species of oak (Quercus spp.), European and American chestnuts (Castanea spp.) plus trees from other botanical families: willows and maples (Salicaceae), redbay (Fagaceae), and alder (Betulaceae).
Pests in Russia that Could Damage North America if Introduced Here
Malus sierversii; photo by Lukacz Szczurowski via Wikimedia
Threat to Apples — Apple Buprestid, Agrilus mali
This Asian beetle has caused extensive mortality of wild apple (Malus sieversii) forests in Xinjiang, China. Wild apple trees are important components of deciduous forests in the Central Asian mountains. The species is also an ancestor of the domestic apple tree. Consequently, the borer is considered a potential threat to cultivated apple trees – presumably everywhere. A. mali might also attack other fruit trees in the Rose family, i.e., Prunus (plums, cherries, peaches, apricots, almonds) and Pyrus (pears).
Unlike most of the other species described here, A. mali is a quarantine pest in Russia and across Europe and the Mediterranean regions – the region where phytosanitary policies are coordinated by the European and Mediterranean Plant Protection Organization (EPPO). Russia bans imports of apple seedlings from infested areas.
China is reported to be experimenting with a possible biocontrol agent, Sclerodermus pupariae (a parasitoid of emerald ash borer).
Threat to Pines and Firs, Already Under Invasive Species Threats
Small Spruce Bark Beetle, Ips amitinus
This European beetle has been considered a secondary pest of dying conifers. Over the last 100 years, it has moved farther North. The first Russian record was 100 years ago, in the region where Russia, Belarus, and Ukraine meet. (Did military action during World War I play a role? This is not discussed by the authors.) By 2022, the beetle occupies 31 million ha. It is probably spread through transport of logs by rail.
In Western Siberia, the spruce beetle has attacked a new host, Siberian pine (Pinus sibirica).
The danger to North America arises from this beetle’s preference for five-needle pines (genus Pinus section Quinquefoliae). North America’s five-needle pines are already under severe pressure from the introduced pathogen white pine blister rust (Cornartium ribicola) and the native mountain pine beetle (Dendroctonus ponderosae).
Four-Eyed Fir Bark Beetle, Polygraphus proximus
This East Asian beetle feeds on firs (Abies spp.). Less commonly, it feeds on other genera in the Pinaceae: spruce (Picea ), pines (Pinus), larch (Larix), hemlock (Tsuga).
This beetle has been spreading west; the first substantiated record in European Russia was 2006 in Moscow. The beetle was probably present in western Siberia in the 1960s, although it was not detected until 2008. Again, the probable pathway of spread is movement of lumber by railroad.
P. proximus vectors an obligate symbiotic fungus, which can rapidly weaken the host. Musolin et al. comment on the beetle’s impacts – which they rarely do in this article. (Does this signify more damaging impacts, or availability of past studies?) They note significant changes in the forests’ ecosystem structure and microclimate, vegetation cover, and local insect fauna.
The danger to North America arises from this beetle’s preference for firs from the sections Balsamea and Grandis. Many North American firs are in these sections, including Fraser fir (Abies fraseri), balsam fir (A. balsamea), subalpine fir (A. lasiocarpa), grand fir (A. grandis), white fir (A. concolor), and others. Several of these firs already are challenged by the introduced balsam woolly adelgid. Firs in central and western Europe are less vulnerable since they are in the section Abies, which the beetle prefers less.
Threats to Poplars
Spotted Poplar Borer,Agrilus fleischeri
This boring beetle is native to northern Asia. It has caused significant mortality in native and exotic Populus plantations in China. Although there have been no reports of this beetle moving beyond its native range, many other Agrilus species have. Canada has twice intercepted adult spotted poplar borers on wood packaging. Musolin et al. fear that the adoption of non-native hosts might trigger an outbreak that would facilitate spread.
Poplar Leafminer, Phyllonorycter populifoliella
balsam poplar; photo by Matt Lavin via Flickr
This micromoth is widely distributed across the Palearctic. It was recently detected on introduced poplars growing in India.
The danger to North America arises from the beetle’s preference for black and balsam poplars. Several species in these taxonomic groups are common in North America, including Populus balsamifera, P. trichocarpa, P. deltoides, and Populus × Canadensis.
Threat to Oaks — Leaf Blotch Miner Moth, Acrocercops brongniardella
This micromoth is widely distributed in Europe and expanding to the north. The pest mines the leaves of several oak species (Quercus spp.), especially English oak, Q. robur; and sometimes European chestnut (Castanea sativa). Leaf blotch miner is considered one of the most important folivore insect pests of oaks in Russia. Damage has been greater in Omsk Oblast (Siberia), where both English oak and the micromoth are introduced species, than in St. Petersburg, which is on the northern limit of their natural range. Musolin et al. fear that the warming climate will lead to the pest causing greater damage in the northern portions of its range.
Threat to Basswood — Lime Leaf Miner, Phyllonorycter issikii
This Asian moth has been moving west since the mid-1980s. It now occupies most of European Russia with some outbreaks in Siberia. In Europe, it is a conspicuous pest of Tilia species.
In these invaded regions, the leaf miner has shifted to novel hosts, including American basswood (T. americana). Basswood is a common plant in the eastern deciduous forest of North America.
Threat to Horse Chestnuts & Urban Trees — Horse-Chestnut Leaf Miner, Cameraria ohridella
This tiny moth was unknown to science before the first recorded outbreak in the late 1980s. Over the next three decades it spread to most of Europe, where horse chestnut (Aesculus hippocastanum)has been widely planted for three centuries. It has caused significant damage.
The first Russian detection was in Kaliningrad, on the shores of the Baltic Sea, in 2003. The leaf miner now occupies 69% of administrative units of European Russia. It is considered one of the Top 100 most dangerous invasive species in Russia.
In North America, the moth might attack native horse chestnuts, Ae. octandra (=flava) and Ae. glabra. Urban plantings are at particular risk because the leaf miner might attack both European horse chestnuts and two non-native maples that have been planted widely, sycamore maple (Acer pseudoplatanus) and Norway maple (A. platanoides). Data cited by Musolin et al. are contradictory regarding larval development on the maples. Once introduced, the leaf miner is difficult to contain because it spreads through natural flight of adults, wind-blown leaves, hitchhiking on vehicles, and movement of infected plants.
Shared Pests
Russia has been invaded by two species that have been introduced in many countries (beyond pine wilt nematode). These two entered the country on plants for planting being imported to landscape venues for the XXII Winter Olympic Games – held in Sochi in 2014.
First to arrive was the Box Tree Moth, Cydalima perspectalis. This East Asian species was first detected outside its native range in Germany in 2006. By 2011 it was widespread in European and Mediterranean countries. In 2021, the boxwood moth was found in North America (first Canada, then the United States). [I discuss the boxwood moth briefly here.]
boxtree moth; photographer unknown
In Russia, box tree moth larvae were first recorded in 2012 on the planting stock of its principal host, Buxus sempervirens. The moth quickly spread around the Black Sea region and to the North Caucasus. It spread farther, too: it reached the Kaliningrad Oblast (southeast coast of the Baltic Sea) in 2020. The main pathway of C. perspectalis invasion was the introduction of infested box-wood planting material.
Further spread of C. perspectalis is likely from Russia into the natural forests across the Caucasus (Transcaucasia) and to countries located further south. This is most distressing because the region has extensive natural forests of Buxus sempervirens. In 2015–2017, C. perspectalis almost completely destroyed the natural boxwood populationsin these regions of Russia and further eastwards in Abkhazia. Boxwood stands in Georgia and northern Iran are already suffering intensive defoliation as the result of infection by two non-native pathogens, Calonectria pseudonaviculata [synonym Cylindrocladium buxicola] and Calonectria henricotiae. Damage to these forests could lead to reductions in soil stability and subsequent declines in water quality and flood protection, changes in forest structure and composition, and declines in Buxus-associated biodiversity (at least 63 species of lichens, fungi, chromista and invertebrates might be obligate). (In December 2022, Iryna Matsiakh presented a compelling overview of threats to these forests in a webinar sponsored by the Horticulture Research Initiative; apparently no recording is available.)
The second global invader to appear was the Brown Marmorated Stink Bug, Halyomorpha halys.
This insect from southeast and east Asia invaded the United States in 1996. The first detection in Europe was in Liechtenstein in 2004. In both cases, it spread quickly across these continents.
Russia’s first detection of stinkbug was in 2014 in parks in Sochi and elsewhere along the Black Sea coast. The spread in Russia appears to have been limited to the Black Sea – Caucasus area.
The brown marmorated stinkbug is highly polyphagous, feeding on more than 300 species of plants. In southern Russia, 107 species have been documented as hosts. At times, stinkbug feeding has caused severe losses in yields of fruit and vegetable crops.
Patterns
Musolin et al. stress the importance of the pest shifting to new hosts–usually from the same or a closely related genus. They cite several examples of these shifts occurring in the pest’s native range, including Agrilus planipennis (from local Asian ash species to introduced North American ash species); Phyllonorycter populifoliella and Agrilus fleischeri (from local poplars to widely cultivated introduced North American poplars and hybrids); Agrilus mali (from cultivated to wild apples).
As I noted above, the introduction and spread pathways are the usual ones: plants for planting (three species) and shipments of logs. There is one indication of wood packaging – Spotted Poplar Borer, Agrilus fleischeri at the Canadian border.
Garbelotto, M., G. Lione, and A.V. Martiniuc. date? The alien invasive forest pathogen Heterobasidion irregulare is replacing the native Heterobasidion annosum. Biological Invasions https://doi.org/10.1007/s10530-022-02775-w
Musolin, D.L.; Kirichenko, N.I.; Karpun, N.N.; Aksenenko, E.V.; Golub, V.B.; Kerchev, I.A.; Mandelshtam, M.Y.; Vasaitis, R.; Volkovitsh, M.G.; Zhuravleva, E.N.; et al. Invasive insect pests of forests and urban trees in Russia: Origin, pathways, damage, and management. Forests 2022, 13, 521.
Tanney, J. Forest Health Challenges Exacerbated by a Changing Climate: Swiss Needle Cast and Sooty Bark Disease in B.C. 65th ANNUAL FOREST PEST MANAGEMENT FORUM (Canada). December 7, 2022.
Tsopelas, P., A. Santini, M.J. Wingfield, and Z.W. de Beer. Canker Stain: A Lethal Disease Destroying Iconic Plane Trees. Plant Disease 2017. 101-645-658 American Phytopathological Society
Spruce budworm (Choristoneura fumiferana); photo by Jerald E. Dewey, USFS; via Bugwood; populations of several forest birds, including Cape May, Tennessee and Bay-Breasted warblers, become more numerous during budworm outbreaks
Now that Drs. Ziska and Aucott have educated us about the strong impact atmospheric CO2 can have on both plants and phytopagous insects, I have asked the experts whether these interactions have been incorporated in the models scientists are using to forecast pest activity in American forests as the climate changes.
The answer is no.
bay-breasted warbler; photograph by Dave Inman at Presque Isle State Park, PA; via Flickr
Dr. Bethany A. Bradley, Co-Director, Northeast Climate Adaptation Science Center at the University of Massachusetts, says empirical models of species range shifts typically only use climate and sometimes other environmental factors (like soils or topography) as predictors of potential geography. Inclusion of demographic processes like how plant growth is affected by more or less water, CO2, competition with other plants etc. would require a lot of data. It is currently impossible since there are tens of thousands of plant species interacting in the forests of eastern North America – and perhaps these factors have been analysed for only a hundred of them.
Mike Aucott points to the same difficulty: inclusion of CO2 in models of the future populations of specific plants would be difficult since the impacts vary from species to species and are compounded by other factors such as soil nitrogen levels, moisture levels, temperature, presence of competing plants, etc.
Regarding insects, Dr. Aucott thinks it is clear that some orders, such as Lepidoptera, don’t fare as well when feeding on plants grown under elevated CO2. He is not aware of efforts to model impacts of high CO2 on specific insects or even orders or feeding guilds.
juniper geometer (inchworm); Dr. Tallamy says inchworms are hairless & good tasting – so sought by birds
Dr. Ziska concurs about the difficulties. Dr. Ziska asks why there is so little funding to study these issues, especially given their probable impact on human food supplies and health – as described in his blog and an opinion piece published in Scientific American two years ago.
I hope that scientists, decision-makers, readers of this blog … maybe even the media! – take into consideration these complexities, even if they cannot be defined.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – [but do not address climate or CO2 aspects] review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Wildfire: one of the widely recognized results of climate change (The Pioneer Fire located in the Boise National Forest near Idaho City, ID began on Jul. 18, 2016 and the cause is under investigation. The Pioneer Fire has consumed 96,469 acres. U.S. Forest Service photo. Original public domain image from Flickr)
A guest blog by Michael Aucott. Mike is a retired research scientist of the NJ Department of Environmental Protection. He has also taught chemistry at the College of New Jersey. He is currently a member of the NJDEP Science Advisory Board Standing Committee on Climate and Atmospheric Sciences, and on the board of directors of the PA/NJ Chapter of the American Chestnut Foundation. If you wish to contact Mike, use the contact button on this website. You MUST include your email address; it is not recorded automatically.
Two major perturbations affect Earth and its living systems, climate change and habitat disruptions. Emerging data show that these are more closely related than previously realized; they are connected by carbon dioxide, CO2.
Climate change basics: the physics
Climate change concerns have focused on the alteration of weather and climate due to the increase in atmospheric concentrations of greenhouse gases, primarily carbon dioxide, CO2. The impact of CO2 on climate has been understood for at least 120 years. In 1896 the Nobel-Prize-winning Swedish chemist Svante Arhennius published calculations demonstrating that human emission of CO2, when combined with the positive feedback effects of water vapor, would warm the Earth (Arhennius, 1896). His equation, ΔF = α ln(C/C0), relates the change in climate “forcing” (the degree to which temperature change is forced) to the ratio of the concentration of CO2 currently in the atmosphere (C) to a previous concentration (C0). This equation is still in use today. Arhennius estimated that a doubling of CO2 would warm the Earth by about 4 degrees C. This estimate is not far off from current estimates based on much more elaborate calculations.
This warming impact is caused by the physics of CO2, water vapor, and other “greenhouse” gases. Infrared radiation causes the CO2, water, and other greenhouse gas molecules to vibrate, leading to the absorption of the energy carried by that radiation. Much of the solar energy coming from the sun is not in the infrared frequency range, so it passes through the atmosphere without being absorbed. However, when this energy is absorbed by the surfaces of the Earth and its biota, and is re-radiated as infrared radiation, it is then absorbed by greenhouse gases, warming the planet.
The amount of water vapor in the atmosphere is directly related to the atmosphere’s temperature: warm air holds more water vapor. Human activity hasn’t directly changed the concentration of water vapor in the atmosphere significantly. But by burning fossil fuels, humans have dramatically increased the atmospheric concentration of CO2 and in so doing, also indirectly increased the concentration of water vapor. Just as Arhennius predicted over 120 years ago, this increase in CO2 is warming the Earth.
Ramifications of this warming include increased heat episodes, the intensification of the hydrological cycle (greater frequency of both heavy precipitation events and of droughts), sea level rise due to the melting of land-based glaciers and the thermal expansion of ocean water, and, almost certainly, intensification of storms and an increase in extreme weather. These climate-warming- based perturbations have the potential to influence the functioning of Earth’s biota in many deleterious ways, and clearly can be associated with the many facets of habitat disruption.
Climate change amplifications: the chemistry
But there’s another aspect of CO2 that may be more important insofar as habitat disruption is concerned and that has been largely ignored: chemistry. CO2 is a trace gas as far as we humans and other animals are concerned, unnoticed by our bodies in normal life. But to plants it is a vital food. It is taken up by plants as an essential input to photosynthesis. In this chemical reaction, using the energy of sunlight, plants combine CO2 and water vapor to make oxygen and carbohydrates, represented with a generic formula of CH2O, according to the equation CO2 + H2O → CH2O + O2. Without this reaction, life as we know it would not exist.
The atmospheric concentration of CO2 has varied over time; some 50 million years ago it was considerably higher than today. However, for at least the last three million years, the concentration of CO2 has been in the range of 280 ppm. Over these millions of years biota have adapted to this concentration. But within the last 300 years, one ten thousandth of this period – a blink of an eye in the geological or evolutionary time scale – the concentration of CO2 has shot up to 420 ppm, a 50% increase, due to humanity’s burning of fossil fuels and forests.
Imagine what might happen to a person who had been on a tight dietary budget for most of his or her life but suddenly got access to 50% more carbohydrates, but no more protein or minerals? We could expect major changes in the metabolism of that person. This dramatic change is what has, in effect, happened to the whole of life on Earth. Our planet’s primary biota, plants, now suddenly have the opportunity to gorge on CO2. But their access to other important substances such as nitrogen has not changed. Evidence is accumulating that CO2 at its elevated level of 420 ppm is not, as has been proclaimed by some, a healthy influence but is instead throwing Earth’s ecosystem into disarray.
Much of the recent experimental evidence on the impacts of enriched atmospheric CO2 has been assembled by Lewis Ziska and presented in his new book, Greenhouse Planet: How Rising CO2 Changes Plants and Life as We Know It (Ziska, 2022; see full citation at the end of the blog). The findings documented in this book reveal a variety of impacts of elevated CO2. These impacts include stimulation of growth of invasive plants, decreases in the nutrient content of major crops, and changes in plants’ production of insecticidal, allergenic, and other compounds. The changing chemistry of plants may be contributing to a major die-off of insects and insect-eating animals including birds. Below are some details.
Habitat Disruptions: Stimulation of Invasive Plants
The generally accepted explanation for why some plants are invasive is that they have been introduced to new regions where their historic predators and parasites aren’t present. Without these drags on their growth, they have flourished. That some alien plants are not browsed by white-tailed deer contributes significantly to their invasiveness in Eastern North America. Other factors are clearly involved as well, including changes in the temperature regime and the availability of water and other resources such as nitrogen.
But elevated CO2 is also a factor. In recent years, techniques for measuring responses of organisms in situ under elevated CO2 conditions have been developed, making possible investigations of the impacts of CO2 concentrations that could exist in the future under otherwise relatively realistic conditions. What the actual atmospheric CO2 concentration will be in 2050 or 2100 is difficult to predict; it depends on what humanity does to control emissions. Various scenarios suggest that levels could exceed 500 ppm by 2050 and might exceed 1000 ppm by 2100 (Tollefson, 2020).
cheatgrass; photo by Jaepil Cho
One study found that the invasive weed Canada thistle, Cirsium arvense, responds more strongly to elevated CO2 than soybean, a crop that it often plagues. In a high CO2 environment, this weed’s root system grows strongly enough to make it significantly more resistant to herbicides (Ziska, et al., 2004) (Ziska, 2010). The highly invasive and dangerously flammable cheatgrass (Bromus tectorum), nicknamed “grassoline” by the U.S. Forest Service because of its propensity to intensify wildfires, also responds strongly to elevated CO2 (Ziska, et al., 2005). Also found to be boosted by enriched CO2 is yellow star-thistle (Centaurea solstitialis), considered one of California’s worst weeds. In one study (Dukes, et al., 2011) this plant grew 600% larger in elevated CO2 relative to ambient, while native plants responded much less strongly or not at all. Japanese honeysuckle, Lonicera japonica, which plagues many areas in the U.S., was found to increase in biomass by 135% at a CO2 concentration of 675 ppm while a similar native plant, coral honeysuckle (Lonicera sempervirens) increased by only 40% (Sasek & Strain, 1991). In a field study also involving Japanese honeysuckle (Belote, et al., 2004), researchers found that its above ground net production (ANPP) approximately tripled under enriched CO2 while other plants in the trial showed showed lesser increases or actual decreases.
Other plants have been found to be selectively encouraged by enriched CO2 including cherry laurel (Prunus laurocerasus), invasive in the Pacific Northwest U.S. and the U.K., (Hattenschwiler & Korner, 2002); dalmation toadflax (Linaria dalmatica), invasive in much of North America (Blumenthal, et al., 2013); honey mesquite (Prosopis glandulosa) , invasive in Australia and parts of Africa (Polley, et al., 1996); and kudzu (Pueraria lobata), which afflicts the Southeast U.S. (Sasek & Strain, 1988). Three plants invasive in China or Southeast Asia, American rope (Mikania micrantha), Creeping oxeye (Wedelia trilobata), and a morning glory species (Ipomoea cairica), were found to produce 70.3% greater biomass when grown at a CO2 concentration of 700 ppm while three corresponding indigenous plants Paederia scandens, Wedelia chinensis and Ipomoea pescaprae, produced only 30.5% more biomass (Song, et al., 2009).
yellow star thistle; photo by Eugene Zelenko
The list goes on of studies showing increased growth of some plants under enriched CO2 conditions. As more in situ investigations are undertaken, it seems likely it will become clearer that today’s enriched level of CO2 is helping some plants to become invasive.
Complexities and contradictory findings exist however. Not all plants are stimulated by enriched CO2. An important difference in the response to higher levels of CO2 is whether a plant has a C3 or a C4 photosynthetic mechanism. C4 plants contain a biochemical pump that concentrates CO2, making them more adapted to low CO2 conditions (Hager, et al., 2016). At current levels of CO2, such plants’ need for CO2 is easily met. C3 plants do not have this CO2 concentrating ability, and so higher levels boost their growth. In a broad meta-analysis of literature, the average response to elevated CO2 of 365 C3 plant species and 37 C4 plant species was noted; the response was significantly increased in C3 species but was unchanged in C4 species (Robinson, et al. 2012). One striking example of such a difference was observed in the field study noted above (Belote, et al., 2004), wherein researchers found that Japanese honeysuckle (a C3 plant) was significantly encouraged by elevated CO2 relative to other plants at the same locale. The same study found that another aggressive invader, Japanese stiltgrass (Microstegium vimineum), a C4 plant, was unaffected or even slightly inhibited relative to competing plants’ growth by elevated CO2.
One finding is widespread; most plants studied under enriched CO2 regimes show an increase in biomass and evince a higher ratio of carbon to nitrogen (C/N ratio) in their tissues and an overall decline in nitrogen concentrations than when grown under ambient conditions. Since nitrogen is a key component of protein, this change can be expected to lead to lowered protein content of critical food crops. Some impacts of this change are already well underway because of today’s elevated CO2 concentration. Changes since 1850 in the C/N ratio and in the estimated protein content of an important plant product, pollen, were discovered in a striking study by Lewis Ziska and colleagues (Ziska, et al., 2016). Using archived museum samples, these researchers determined the nitrogen content of pollen of Solidago canadensis (Canada goldenrod) going back to the 1850s. They estimated that the protein content of goldenrod pollen, a vital nutrient for North American bees, has declined in inverse proportion to the rise in atmospheric CO2, dropping from a concentration of approximately 18% in the mid-1800s to approximately 12% today. They pointed out that it is possible that bees are now unable to provide sufficient protein and other nutrients to larvae, and that one of the main reasons for bee declines is malnutrition caused by enriched atmospheric CO2. Other studies also indicate that elevated CO2 could cause lower nitrogen concentrations in plants and lead to less proteinaceous plant parts, including pollen, being available to plant-feeding insects (Hall, et al., 2005; Knepp, et al., 2007).
bumblebee on goldenrod; photo by Keila
The changing C/N ratio is almost certainly already affecting the human food supply. As documented in an extensive review of published findings (Soares, et al., 2019), elevated CO2 has a considerable impact on the accumulation of minerals and protein in plants, with many plant species showing declines in both quality and quantity of key nutrients. These changes have worrisome implications for human nutrition and may already be responsible for increasing incidences of dietary deficiency in some areas. Lewis Ziska discusses the likely impact of rising CO2 on the future human food supply in his recent post. A number of studies showing declines in protein and also other nutrients such as zinc in food crops important to humanity are also described in Ziska’s new book, Greenhouse Planet, noted above.
Habitat Disruptions: Other Changes in Plant Chemistry
Other changes in plants besides nutritional content may be driven by enriched CO2. Plants produce a variety of secondary metabolites. Most plants use the C3 mechanism; with 50% more available of a key input, some changes in these plants’ production of such chemicals can be expected. Some changes have been observed. Mohan et al. (2006) report that enriched CO2 in an intact forest system increased water use efficiency, growth, and population biomass of poison ivy (Toxicodendron radicans) and that high-CO2 plants also produced a more toxic form of the allergenic compound urushiol.
Quercus chapmanii; photo by Mary Keim at Seminole State Forest, Florida
Many of the phytochemicals plants produce function as defenses against insect predation, and changes in such production have been found to impact herbivore feeding. For example, Landosky and Karowe (2014) suggest that specialist herbivores may have to contend with more effective chemical defenses by plants under elevated CO2. Hall, et al. (2005), in a study involving several oak and one legume species in a scrub oak ecosystem in Florida (see photo above), found that 700 ppm CO2 levels led to decreased damage to plants by four of six insect groups investigated. They did not see increases in plants’ production of carbon-based secondary metabolites, including total phenolic compounds, condensed tannins, hydrolyzable tannins, cellulose, hemicellulose, and lignin however. They concluded that the primary driver of decreased insect predation under elevated CO2 was lower overall plant nitrogen levels. They stated that the decline of nitrogen levels in foliage under elevated CO2 indicated lower foliar quality and hypothesized that the reductions in insect feeding stemmed from the combined effects of nutrient limitation and increases in parasitism and predation on the nutrient-constrained insects. They further stated that although insects try to compensate for lower nutrient content of leaves by eating more, they did not see an increased portion of damaged leaves in their study. These researchers did not report measurements of terpenoid compounds however, which are reported to represent the largest class of secondary metabolites (Wikipedia, 2022). In another study (Hall, et al., 2005a) found that concentrations of condensed tannins were higher in oak leaf litter under elevated CO2, which suggests that enhanced production of insecticidal compounds or other changes to plant tissues could affect not only insects that consume living plant tissue, but also detritivores.
Robinson et al. (2012) also investigated plants’ production of secondary metabolites in their literature review. Looking at all plant groups, they found that under elevated CO2 the production of nitrogen-based secondary metabolites (e.g., alkaloids, cyanogenic glycosides, and glucosinolates) decreased by 16% while the carbon-based secondary metabolites total phenolics, condensed tannins, and flavonoids increased by 19%, 22%, and 27% respectively. Another carbon-based metabolite, terpenoids, declined by 13%. They further divided plants into grasses, shrubs, herbs/forbs, and trees and found differing responses to elevated CO2. Trees, for example, showed increased production of total glycosides and total phenolics, little change in production of total flavonoids, and a decline in the production of total terpenes. Like Hall et al., (2005), Robinson et al. found a strong and significant decrease in nitrogen concentrations under elevated CO2 for C3 plants. A decrease did not show up for C4 plants.
In addition to chemical defenses, plants have physical characteristics such as surface waxes, trichomes, secretory canals, and tissue toughness-enhancing substances such as lignin. All of these features can reduce the edibility of plants for arthropod herbivores. Robinson et al. (2012) found consistent responses to these characteristics under elevated CO2; leaf toughness and specific leaf weight increased by 11% and 18%, respectively, while specific leaf area did not show a significant change. They concluded that there is an increase in general “toughness” of leaves under elevated CO2. As did Hall et al., (2005), Robinson et al. concluded that elevated CO2 will induce changes in plant chemistry, physiology, and morphology that are likely to impact the nutritional quality of host plants for insect herbivores.
Habitat Disruptions: Changes in Plant Chemistry and Insect Decline
Numerous studies have documented a recent and dramatic decline in insect populations and discussed the probable cascading impacts of such declines through the food chain, affecting spiders, lizards, birds, and other organisms (Samways, et al., 2020; Cardoso, et al., 2020; Sánchez-Bayoa & Wyckhuys, 2019; Tallamy & Shriver, 2021). It has been argued that the main drivers of insect species declines are habitat loss and conversion to intensive agriculture and urbanization; pollution, mainly by synthetic pesticides and fertilizers; biological factors, including pathogens and introduced species; and climate change.
But a puzzling aspect is that some insect declines have been observed in nature preserves (Vogel, 2017) that presumably are not greatly affected by most of the above drivers. One example is a study spanning 27 years that found a 76% decline in flying insect biomass at several of Germany’s protected areas subject to rather low levels of human disturbance (Hallmann et al., 2017). Another study in rainforests of Puerto Rico, apparently not subject to influences such as light pollution, habitat loss, pesticides, or agriculture, reported biomass losses between 98% and 78% for ground-foraging and canopy-dwelling arthropods over a 36-year period, (Lister and Garcia, 2018). This leaves climate change as the likely culprit. But although the varied impacts of climate change, including heat episodes, drought, and other episodes of extreme weather could impact insect populations in remote as well as populated areas, the trends observed appear to far exceed the magnitude of such climate-related disturbances over the last several decades.
tent caterpillars; Shiela Brown, Public Domain Pics
Another puzzling aspect is that not all insect orders or feeding guilds seem to be equally affected. Sanchez-Bayoa & Wychuys (2019) whose article reports on a review of 73 historical reports, state that Lepidoptera, Hymenoptera and dung beetles (Coleoptera) appear to be the taxa most affected in terrestrial environments. Robinson et al. (2012) found that phloem feeders such as Homoptera respond positively to elevated CO2 while foliage feeders/Lepidoptera respond negatively. Lepidoptera were among the most impacted; relative growth rate, fecundity, and abundance all declined under high CO2 conditions, while relative consumption rate, total consumption, and development time all increased.
Most Lepidoptera are herbivorous, feeding in their larval stage, caterpillars, on plants. Caterpillars are key components of the terrestrial ecology; in most forests of the world, caterpillars consume more living leaves than all other animals combined (Janzen, 1988). Insect herbivores such as caterpillars are near the hub of most terrestrial food webs, comprising essential food for insect predators and parasitoids, spiders, amphibians, lizards, rodents, bats, birds, and even higher predators such as foxes and bears (Burghardt et al., 2010). At least 310 North American bird species are known to feed extensively on caterpillars, and the majority of terrestrial birds rely on insects during reproduction and other nutrient-limited periods in the annual cycle (Narango, Tallamy & Marra, 2018). Caterpillars and moths are the largest component of nestling diets in hundreds of species of migrant bird species (Tallamy & Shriver, 2021); they are among the “little things that run the world” (Wilson, 1987).
Carolina chickadee; one of the birds Dr. Tallamy focuses on because it feeds its young on caterpillars; photo by Dan Pancamo; through Wikimedia
Habitat Disruptions and Climate Change: Connected by CO2
The apparently heightened rate of decline of insect herbivores such as caterpillars compared to some other insects, and the findings that many declines have been observed in areas relatively unimpacted by direct human influences such as light pollution, pesticides, and land-use change, point to the likelihood of a broad, perhaps ubiquitous, cause. Climate change is such a broad cause. Even broader and more ubiquitous is the main driver of climate change, CO2. Every plant in the world is constantly bathed in an enriched concentration of this gas. A conclusion seems likely: CO2 is not only causing global warming and climate change but is also affecting life on this planet in ways that, while still poorly understood, are already reducing the nutritive value of food crops, may be a significant cause of the spread of invasive plants, and may be the main driver of insect declines and the cascading impacts of such declines on insect-eating animals such as birds.
What to do?
To mitigate climate change and, as argued here, to mitigate habitat disruption, the steady rise in the atmosphere’s burden of CO2 must be halted, and then steps must be taken to lower the current concentration to a healthier level. These are not hopeless tasks. Although what has been a relentless rise in CO2 emissions at the global level continues, increases have slowed and even stopped in some parts of the world. Accelerating the development of low- and zero-carbon energy sources and encouraging energy conservation, as will be done through the U.S.’s Inflation Reduction Act, will further this progress.
More will be needed. Putting a significant and steadily increasing price on the carbon in fossil fuels is arguably the most important next step. Fossil fuels enjoy a free ride. The byproduct of their combustion, CO2, is dumped with little or no restrictions into the world’s atmosphere. A price on carbon would end this inequity. There are ways this could be done in a revenue-neutral (“fee and rebate”) manner that would avoid harm to economies and those with low- and moderate-incomes. A major step forward in pricing carbon by the European Union, a carbon border adjustment mechanism, is close to implementation. For more on this and other developments in cutting CO2 emissions, see the analyses and insights of the Carbon Tax Center and learn more about actions you can take to influence governments with Citizens’ Climate Lobby.
Not discussed here, but another stark example of habitat disruption is the increasing acidification of the world’s oceans caused by the dissolution of atmospheric CO2 in the waters. The ocean’s average pH has dropped from 8.2 to 8.1 within the last 200 years. Because pH is a logarithmic scale, this represents an increase in hydrogen ion concentration of over 25%, a change that is already threatening some marine creatures. More on this is available from many sources; e.g., Kolbert (2014).
References
Arhennius, Svante, 1896, On the Influence of Carbonic Acid in the Air upon the Temperature on the Ground, Philosophical Magazine and Journal of Science, 41, 237-276.
Belote, R., J. Weltzin, and R. Norby, 2004, Response of an Understory Plant Community to Elevated [CO2] Depends on Differential Responses of Dominant Invasive Species and Is Mediated by Soil Water Availability, New Phytologist 161, 827-835.
Blumenthal, D., V. Resco, J. Morgan, D. Williams, D. LeCain, E. Hardy, E. Pendall, and E. Bladyka, 2013, Invasive Forb Benefits from Water Savings by Native Plants and Carbon Fertilization Under Elevated CO2 and Warming, New Phytologist 200, 1156-1165.
Burghardt, Karin T., D. W. Tallamy, C. Philips, and K. Shropshire, 2010, Non-native plants reduce abundance, richness, and host specialization in lepidopteran communities, Ecosphere 1: 1-22.
Dukes, J., N. Chiariello, S. Loarie, and C. Field, 2011, Strong Response of an Invasive Plant Species (Centaurea solstitialis L.) to Global Environmental Changes, Ecological Applications 21, 1887-1894.
Hall, M., P. Stiling, B. Hungate, B. Drake, and M. Hunter, 2005a, Effects of elevated CO2 and herbivore damage on litter quality in a scrub oak ecosystem, Journal of Chemical Ecology, 31, 2343-2356.
Hall, M., P. Stiling, D. Moon, B. Drake, and M. Hunter, 2005, Effects of elevated CO2 of foliar quality and herbivore damage in a scrub oak ecosystem. Journal of Chemical Ecology 31, 267-286.
Hallmann, C.A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., Stenmans, W., Müller, A., Sumser, H., Hörren, T., Goulson, D., de Kroon, H., 2017, More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 12, e0185809.
Hattenschwiler, S. and C. Korner, 2003, Does Elevated CO2 Facilitate Naturalization of the Non-indegenous Prunus laurocerasus in Swiss Temperate Forests?, Functional Ecology 17, 778-785.
Janzen, Daniel H., 1988, Ecological Characterization of a Costa Rican Dry Forest Caterpillar Fauna, Biotropica, 20, 120-135.
Knepp, R., J. Hamilton, A. Zangeri, M. Berenbaum, and E. Delucia, 2007, Foliage of oaks grown under elevated CO2 reduces performance of Antherae Polyphemus (Lepidoptera: Saturnidae), Environmental Entomology 36, 609-617.
Kolbert, E., 2014, The Sixth Extinction, Henry Holt & Co., NY
Landosky, J., and D. Karowe, 2014, Will chemical defenses become more effective against specialist herbivores under elevated CO2? Global Change Biology, 20, 3159–3176.
Lister, B., and A. Garcia, 2018, Climate-driven declines in arthropod abundance restructure a rainforest food web, PNAS 115, E10397–E10406
Mohan, J., L. Ziska, W. Schlesinger, R. Thomas, R. Sicher, K. George, and J. Clark, 2006, Biomass and toxicity responses of poison ivy (Toxicodendron radicans) to elevated atmospheric CO2. PNAS 103, 9086-9089.
Narango, D., D. Tallamy, and P. Marra, 2018, Nonnative plants reduce population growth of an insectivorous bird, PNAS 115: 11549–11554.
Polley, H., H. Johnson, H. Mayeux, C. Tischler, and D. Brown, 1996, Carbon Dioxide Enrichment Improves Growth, Water Relations, and Survival of Droughted Honey Mesquite (Prosopis glandulosa) Seedlings, Tree Physiology, 16, 817-823.
Robinson, E., G. Ryan, and J. Newman, 2012, A meta-analytical review of the effects of elevated CO2 on plant-arthropod interactions highlights the importance of interacting environmental and biological variables, New Phytologist 194, 321-336.
Sánchez-Bayoa, F. and K. Wyckhuys, 2019, Worldwide decline of the entomofauna: A review of its drivers, Biological Conservation 232, 8-27.
Sasek, T. and B. Strain, 1988, Effects of Carbon Dioxide Enrichment on the Growth and Morphology of Kudzu (Puerarialobata), Weed Science 36, 28 – 36, DOI: https://doi.org/10.1017/S0043174500074415
Sasek, T. and B. Strain, 1991, Effects of CO2 Enrichment on the Growth and Morphology of a Native and Introduced Honeysuckle Vine, American Journal of Botany 78, 69-75.
Soares, J., C. Santos, S Carvalho, M Pintado, and M. Vasconceios, 2019, Preserving the nutritional quality of crop plants under a changing climate: importance and strategies. Plant and Soil 443, 1-26. https://doi.org/10.1007/s11104-019-04229-0
Song, L., J. Wu, C. Li, F. Li, S. Peng, and B. Chen, 2009, Different responses of invasive and native species to elevated CO2 concentration, Acta Oecologica 35, 128-135.
Tallamy, D. and W.G. Shriver, 2021, Are declines in insects and insectivorous birds related? Ornithological Applications 123: 1–8, DOI: 10.1093/ornithapp/duaa059
Vogel, G., 2017, Where Have All the Insects Gone? Surveys in German nature preserves point to a dramatic decline in insect biomass. Key members of ecosystems may be slipping away, Science 356, 376-379.
Wilson, Edward O., 1987, The Little Things That Run the World (The Importance and Conservation of Invertebrates), Conservation Biology, 1, 344-346.
Ziska, L., 2010, Elevated Carbon Dioxide Alters Chemical Management of Canada Thistle in No-till Soybean, Field Crops Research 119, 299-303.
Ziska, L., Shaun Falukner, and John Lydon, 2004, Changes in biomass and root: Shoot Ratio of Field-grown Canada Thistle (Cirsium arvense), a Noxious, Invasive Wed, with Elevated CO2: Implications for Control with Glyphosate, Weed Science 52, 584-588.
Ziska, L.H., J.S. Pettis, J. Edwards, J.E. Hancock, M.B. Tomecek, A. Clark, J.S. Dukes, I. Loladze, and H.W. Polley, 2016, Rising atmospheric CO2 is reducing the protein concentration of a floral pollen source essential for North American bees, Proc. R. Soc. B, 283, 20160414, http://dx.doi.org/10.1098/rspb.2016.0414
Ziska, Lewis, 2022, Greenhouse Planet: How Rising CO2 Changes Plants and Life as We Know It, Columbia University Press, NY.