emerald ash borer; some of PPA grants are funding evaluation of biocontrol efficacy
USDA APHIS has released information about its most recent annual allocation of funds under the Plant Pest and Disease Management & Disaster Prevention Program under §7721 of the Plant Protection Act. (Also see Fading Forests II and III; links provided at the end of this blog.) These funds support both critical needs and opportunities to strengthen the nation’s infrastructure for pest detection, surveillance, identification, and threat mitigation. Since 2009, this USDA program has provided nearly $940 million to more than 5,890 projects.
For FY25 APHIS allocated $62.725 million to fund 339 projects, about 58% of the proposals submitted. About $10 million has reserved for responding to pest and plant health emergencies throughout the year.
According to APHIS’ press release, the highest amount of funds (almost $16 million) is allocated to the category “Enhanced Plant Pest/Disease Survey.” Projects on “Enhanced Mitigation Capabilities” received $13.6 million. “Targetting Domestic Inspection Efforts to Vulnerable Points” received nearly $6 million. “Improving Pest Identification and Detection Technology” was funded at $5 million. Outreach & education received $4 million. I am not sure why these do not total $63 million.
Funding for States and Specific Pests
Wood-boring insects received about $2.3 million. These included more than $869,800 to assess the efficacy of biocontrol for controlling emerald ash borer (EAB) Agrilus planipennis, $687,410 was provided for various detection projects, and $450,000 for outreach efforts related to various pests. Ohio State received $93,000 to optimize traps for the detection of non-native scolytines (bark beetles).
Biocontrol efficacy will also be assessed for hemlock woolly adelgid, invasive shot hole borers, cactus moth, and several invasive plants (including Brazilian pepper). (Contact me to obtain a copy of CISP’s comments on this biocontrol program.)
Opuntia basilaris in Anza Boreggo; one of flat-padded Opuntia vulnerable to the cactus moth; photo by F.T. Campbell
Funding for other pests exceeded $1 million for spotted lanternfly (nearly $1.4 million), Asian defoliators ($1.2 million) and box tree moth (just over $1 million).
$630,000 was provided for detection surveys and studies of the sudden oak death pathogen Phytophthora ramorum, especially how it infects nursery stock. Nursery surveys are funded in Alabama, Louisiana, North Carolina, Ohio, Oklahoma, Pennsylvania, South Carolina, Tennessee, Virginia, and West Virginia. Most of these states are in regions considered most at risk to SOD infection of wildland plants.
sudden oak mortality of tanoak trees in southern Oregon; photo by Oregon Department of Forestry
Oregon received much-deserved $41,000 to evaluate the threat of the NA2 and EU2 lineages of P. ramorum to nurseries and forests Oregon also received $104,000 to respond to the detection of Phytophthora austrocedri in nurseries in the state. The Oregon outbreak has been traced to Ohio, but I see no record of funds to assist that state in determining how it was introduced.
Asian defoliator (e.g., Lymantrid moths) surveys have been funded for several years. This year’s projects are in Alaska, Arkansas, California, Kentucky, Maryland, Massachusetts, Mississippi, Montana, Nevada, North Carolina, Oregon, Tennessee, Texas, Washington, and West Virginia. While I agree that the introduction risk is not limited to coastal states with maritime ports, I don’t what criteria were applied in choosing the non-coastal states which are funded to search for these insects
Spotted lanternfly surveys (including technological improvements) or related outreach are funded in Alabama, Connecticut, Delaware, Kentucky, New Hampshire, New Jersey, North Carolina, Oregon, Pennsylvania, and Tennessee. California’s project is focused on postharvest treatments.
The Don’t Move Firewood project continues to be funded by APHIS. Several states also direct attention specifically to the firewood pathway: Kentucky, Maine, and Michigan.
I applaud the precautionary funding of the Agriculture Research Service to generate of high-quality genomic resources for managing the causal agent of Japanese oak wilt Dryadomyces quercivorous
Florida Department of Agriculture, North Carolina State University, and West Virginia University each received more than $100,000 to improve detection and management of invasive hornets.
Tennessee State University got $100,000 to continue efforts to detect and understand Vascular Streak Dieback in redbud Cercis canadensis.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
obvious risk of pest introduction! photo by F.T. Campbell
Because of the many damaging insects introduced in wood packaging, I often blog about numbers of shipping containers entering the country. [On the “nivemnic.us” website, scroll down below “archives” to “categories”, then click on “wood packaging” to see my previous blogs discussing this issue.]
The Department of Homeland Security’s Bureau of Customs and Border Protection (CBP) reports processing 36.6 million shipping containers holding imports in Fiscal Year 2023 – which ended in September 2023. These presumably included about 13 – 16 million containers arriving via ship from Asia, Europe, and other overseas trading partners. The remaining millions probably entering from Mexico and Canada via land transport. Together, Mexico and Canada provided 30% of U.S. imports in 2022.
It is difficult to pin down the actual number of containers entering the country. In contrast to the figure provided by CBP, Laura Robb of the Journal of Commerce reports that 25.6 million TEUs carrying imports entered the country in 2024. This figure apparently includes containers carried by all forms of transport. CBP counts containers by actual numbers, and about 90% of waterborne containers are actually 40 feet long, not the 20 feet measured by “TEU” (U.S. DoT). Halving the JOC number results in a total of about 13 million – well below that reported by CBP.
Overall volumes of imports carried by ship continue to rise. The monetary value of goods imported by the U.S. in maritime trade grew 15% from 2021 to 2022 (U.S. DoT). Robb reported that trade experts believe imports rose another 15% between 2023 and 2024. This rise is driven by retailers trying to protect themselves from a possible longshoremen’s strike (which might occur beginning 15 January), Trump’s threatened tariffs (he might act as early as 20 January); and the annual slowdown of production in Asia during Tet (which begins on 29 January). If import volumes meet expectations and continue through April, the series will outdo the previous (pandemic-era) record of 19 straight months when imports exceeded 2 million TEUs. What happens later in 2025 depends in part on whether the anticipated strike happens and/or actual levels of any new tariffs.
One concern about imports from Mexico and Canada is that some proportion of these goods actually originated in Asia or Europe, but were shipped through Mexican or Canadian ports. I have not found a source to clarify how many shipments fit this pattern. USDA APHIS used to blame forest pests introduced to the Great Lakes region on goods transported from the principal Canadian Atlantic port, St. John, Nova Scotia.
A useful publication for identifying where the pest-introduction risk is highest are the annual reports issued by U.S. Department of Transportation’s Bureau of Transportation Statistics. In calendar year 2022, U.S. maritime ports handled just under 43% of U.S. international trade (measured by value). There are two caveats: the data include both imports and exports; and the most recent data are from 2021.
Two-thirds of America’s maritime cargo (imports and exports) is shipped in traditional containers. This includes most consumer goods. The top 25 container ports handled a total of 45.6 million TEU (U.S. DoT). Map 4-3 in the report shows these ports and the proportions that are imports and exports.
The highest-ranking Container Ports in 2021 are those we expect. The ports of Los Angeles and Long Beach were numbers one and two. Together they received 10.7 million TEU. The third highest number of containers entered through the Port of New York & New Jersey. Nearly 5 million TEU entered there. The Port of Savannah ranked fourth. Savannah and nearby Charleston (ranked seventh) handled 4.2 million incoming TEUs in 2021.
Ranked above Charleston were the Port of Virginia and Houston. Each processed approximately 1.8 million containers filled with imports. Three West coast ports follow: Oakland, California and Tacoma and Seattle. Just over 1 million TEUs entered Oakland. The two Washington ports received a little over 1.5 million. Florida has four ports ranked in the “top 25”. In total, they processed 1.2 million TEU; most entered through PortMiami and Port Everglades. Baltimore, Philadelphia, Mobile, New Orleans, Wilmington, North Carolina and Wilmington, Delaware, South Jersey Port Corporation, and Boston all handled less than 500 imported containers in 2021. Domestic shipments from other U.S. states dominated containers processed through the ports of San Juan, Honolulu, and Alaska.
gantry crane in operation at the Port of Savannah; photo by F.T. Campbell
The top ports must have appropriate facilities needed to load / unload container vessels efficiently– that is, adequate numbers of gantry cranes, especially super post-Panamax cranes, which have the greatest capacity. The top 25 container ports of 2021 operated a total of 539 ship-to-shore gantry cranes in 2023, of which 322 (60%) are post-Panamax cranes. Ports are adding cranes – there were 29 more in 2023 than in 2021. The Port of Virginia appears to be striving for significant increases in tonnage; it has 28 Panamax cranes, more than Charleston and almost as many as Savannah (U.S. DoT).
Another important port component is efficient facilities to load containers onto rail cars or trucks for transfer to land-based warehouses and retailers. Ports have more than one terminal; for example, the Port of Long Beach has six, New York/New Jersey has five. Nationwide, 70% of container terminals have on-dock facilities to transfer containers directly onto rail cars. All but three of the 33 terminals located at Long Beach. Los Angeles, New York, Savannah, Charleston, Houston 2/2, Seattle, and Tacoma have on-dock transfer equipment.
The U.S. DoT reports also inform us about the top 25 ports that handle other categories of cargo: overall tonnage, dry and liquid bulk cargo, break bulk cargo, and roll-on-roll-off cargo. Visit the report to view these data.
SOURCES
Robb, L. 2024. U.S. import “surge” to persist into spring amid continued frontloading: retailers. Journal of Commerce Daily Newswire December 10, 2024
U.S. Department of Transportation, Bureau of Transportation Statistics, Annual Report 2024 Port Performance Freight Statistics January 2024 https://www.bts.gov/explore-topics-and-geography/modes/maritime-and-inland-waterways/2024-port-performance-freight
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
ʻŌhiʻa trees killed by ROD; photo by Richard Sniezko, USFS
Several Hawaiian tree species are at risk due to introduced forest pests. Two of the Islands’ most widespread species are among the at-risk taxa. Their continuing loss would expose watersheds on which human life and agriculture depend. Habitats for hundreds of other species – many endemic and already endangered – would lose their foundations. These trees also are of the greatest cultural importance to Native Hawaiians.
I am pleased to report that Hawaiian scientists and conservationists are trying to protect and restore them.
Other tree species enjoy less recognition … and efforts to protect them have struggled to obtain support.
1) koa (Acacia koa)
Koa is both a dominant canopy tree and the second-most abundant native tree species in Hawai`i in terms of areas covered. The species is endemic to the Hawaiian archipelago. Koa forests provide habitat for 30 of the islands’ remaining 35 native bird species, many of which are listed under the U.S. Endangered Species Act. Also dependent on koa forests are native plant and invertebrate species and the Islands’ only native terrestrial mammal, the Hawaiian hoary bat. Finally, koa forests protect watersheds, add nitrogen to degraded soils, and store carbon [Inman-Narahari et al.]
Koa forests once ranged from near sea level to above 7000 ft (2100 m) on both the wet and dry sides of all the large Hawaiian Islands. Conversion of forests to livestock grazing and row-crop agriculture has reduced koa’s range. Significant koa forests are now found on four islands – Hawai’i, Maui, O‘ahu, and Kauaʻi. More than 90% of the remaining koa forests occur on Hawai`i Island (the “Big Island) [Inman-Narahari et al.]
In addition to its fundamental environmental role, koa has immense cultural importance. Koa represents strength and the warrior spirit. The wood was used traditionally to make sea-going canoes. Now Koa is widely used for making musical instruments, especially guitars and ukuleles; furniture, surfboards, ornaments, and art [Inman-Narahari et al.]
Koa timber has the highest monetary value of any wood harvested on the Islands. However, supplies of commercial-quality trees are very limited (Dudley et al. 2020). Harvesting is entirely from old-growth forests on private land. [Inman-Narahari et al.]
Koa forests are under threat by a vascular wilt disease caused by Fusarium oxysporum f. sp. koae (FOXY). This disease can kill up to 90% of young trees and – sometimes — mature trees in native forests. The fungus is a soil-dwelling organism that spreads in soil and infects susceptible plants through the root system (Dudley et al. 2020).
Conservation and commercial considerations have converged to prompt efforts to breed koa resistant to FOXY. Conservationists hope to restore native forests on large areas where agriculture has declined. The forestry industry seeks to enhance supplies of the Islands’ most valuable wood. Finally, science indicated that a breeding program would probably be successful. Field trials in the 1990s demonstrated great differences in wilt-disease mortality among seed sources (the proportion of seedlings surviving inoculation ranged from 4% to 91.6%) [Sniezko 2003; Dudley et al. 2009].
In 2003, Dudley and Sniezko outlined a long-term strategy for exploring and utilizing genetic resistance in koa. Since then, a team of scientists and foresters has implemented different phases of the strategy and refined it further (Dudley et al. 2012, 2015, 2017; Sniezko et al. 2016]
First, scientists determined that the wilt disease is established on the four main islands. Having obtained more than 500 isolates of the pathogen from 386 trees sampled at 46 sites, scientists tested more than 700 koa families from 11 ecoregions for resistance against ten of the most highly virulent isolates (Dudley et al. 2020).
The Hawaiian Agricultural Research Center (HARC), supported by public and private partners, has converted the field-testing facilities on Hawai`i, Maui, and Oahu into seed orchards. The best-performing tree families are being grown to maturity to produce seeds for planting. It is essential that the seedlings be not just resistant to FOXY but also adapted to the ecological conditions of the specific site where they are to be planted [Dudley et al. 2020; Inman-Narahari et al. ] Locally adapted, wilt-resistant seed has been planted on Kauaʻi and Hawai`i. Preparations are being made to plant seed on Maui and O‘ahu also. Scientists are also exploring methods to scale up planting in both restoration and commercial forests [R. Hauff pers. comm.].
koa; photo by David Eickhoff via Flickr
Restoration of koa on the approximately half of lands in the species’ former range that are privately owned will require that the trees provide superior timber. Private landowners might also need financial incentives since the rotation time for a koa plantation is thought to be 30-80 years. [Inman-Narahari et al.]
Plantings on both private and public lands will need to be protected from grazing by feral ungulates and encroachment by competing plants. These management actions are intensive, expensive, and must be maintained for years.
Some additional challenges are scientific: uncertainties about appropriate seed zones, efficacy of silvicultural approaches to managing the disease, and whether koa can be managed for sustainable harvests. Human considerations are also important: Hawai`i lacks sufficient professional tree improvement or silvicultural personnel, a functioning seed distribution and banking network — and supporting resources. Finally, some segments of the public oppose ungulate control programs. Inman-Narahari et al.
Finally, scientists must monitor seed orchards and field plantings for any signs of maladaptation to climate change. (Dudley et al. 2020).
2) ʻŌhiʻa Metrosideros polymorpha)
ʻŌhiʻa lehua is the most widespread tree on the Islands. It dominates approximately 80% the biomass of Hawaii’s remaining native forest, in both wet and dry habitats. ʻŌhiʻa illustrates adaptive radiation and appears to be undergoing incipient speciation. The multitude of ecological niches and their isolation on the separate islands has resulted in five recognized species in the genus Metrosideros. Even the species found throughout the state, Metrosideros polymorpha, has eight recognized varieties (Luiz et al. (2023) (some authorities say there are more).
Loss of this iconic species could result in significant changes to the structure, composition, and potentially, the function, of forests on a landscape level. High elevation ‘ohi‘a forests protect watersheds across the state. ʻŌhiʻa forests shelter the Islands’ one native terrestrial mammal (Hawaiian hoary bat), 30 species of forest birds, and more than 500 endemic arthropod species. Many species in all these taxa are endangered or threatened (Luiz et al. 2023). The increased light penetrating interior forests following canopy dieback facilitates invasion by light-loving non-native plant species, of which Hawai`i has dozens. There is perhaps no other species in the United States that supports more endangered taxa or that plays such a geographical dominant ecological keystone role [Luiz et al. 2023]
For many Native Hawaiians, ‘ōhi‘a is a physical manifestation of multiple Hawaiian deities and the subject of many Hawaiian proverbs, chants, and stories; and foundational to the scared practice of many hula. The wood has numerous uses. Flowers, shoots, and aerial roots are used medicinally and for making lei. The importance of the biocultural link between ‘ōhi‘a and the people of Hawai`i is described by Loope and LaRosa (2008) and Luiz et al. (2023).
In 2010 scientists detected rapid mortality affecting ‘ōhi‘a on Hawai‘i Island. Scientists determined that the disease is caused by two recently-described pathogenic fungi, Ceratocystis lukuohia and Ceratocystis huliohia. The two diseases, Ceratocystis wilt and Ceratocystis canker of ʻōhiʻa, are jointly called “rapid ‘ōhi‘a death”, or ROD. The more virulent species, C. lukuohia, has since spread across Hawai`i Island and been detected on Kaua‘i. The less virulent C. huliohia is established on Hawai`i and Kaua‘i and in about a dozen trees on O‘ahu. One tree on Maui was infected; it was destroyed, and no new infection has been detected [M. Hughes pers. comm.] As of 2023, significant mortality has occurred on more than one third of the vulnerable forest on Hawai`i Island, although mortality is patchy.
[ʻŌhiʻa is also facing a separate disease called myrtle rust caused by the fungus Austropuccinia psidii; to date this rust has caused less virulent infections on ‘ōhi‘a.]
rust-killed ‘ōhi‘a in 2016; photo by J.B. Friday
Because of the ecological importance of ‘ōhi‘a and the rapid spread of these lethal diseases, research into possible resistance to the more virulent pathogen, C. lukiohia began fairly quickly, in 2016. Some ‘ōhi‘a survive in forests on the Big Island in the presence of ROD, raising hopes that some trees might possess natural resistance. Scientists are collecting germplasm from these lightly impacted stands near high-mortality stands (Luiz et al. 2023). Five seedlings representing four varieties of M. polymorpha that survived several years’ exposure to the disease are being used to produce rooted cuttings and seeds for further evaluation of these genotypes.
ʻŌhiʻa flowers
Encouraged by these developments, and recognizing the scope of additional work needed, in 2018 stakeholders created a collaborative partnership that includes state, federal, and non-profit agencies and entities, ʻŌhiʻa Disease Resistance Program (‘ODRP) (Luiz et al. 2023). The partnership seeks to provide baseline information on genetic resistance present in all Hawaiian taxa in the genus Metrosideros. It aims further to develop sources of ROD-resistant germplasm for restoration intended to serve several purposes: cultural plantings, landscaping, and ecological restoration. ‘ODRP is pursuing screenings of seedlings and rooted cuttings sampled from native Metrosideros throughout Hawai`i while trying to improve screening and growing methods. Progress will depend on expanding these efforts to include field trials; research into environmental and genetic drivers of susceptibility and resistance; developing remote sensing and molecular methods to rapidly detect ROD-resistant individuals; and support already ongoing Metrosideros conservation. If levels of resistance in wild populations prove to be insufficient, the program will also undertake breeding (Luiz et al. 2023).
To be successful, ‘ODRP must surmount several challenges (Luiz et al. 2022):
increase capacity to screen seedlings from several hundred plants per year to several thousand;
optimize artificial inoculation methodologies;
determine the effects of temperature and season on infection rates and disease progression;
find ways to speed up seedlings’ attaining sufficient size for testing;
develop improved ways to propagate ʻōhiʻa from seed and rooted cuttings;
establish sites for field testing of putatively resistant trees across a wide range of climatic and edaphic conditions;
establish seed orchard, preferably on several islands;
establish systems for seed collection from the wide variety of subspecies/varieties;
if breeding to enhance resistance is appropriate, it will be useful to develop high-throughput phenotyping of the seed orchard plantings.
Developing ROD-resistant ‘ōhi‘a is only one part of a holistic conservation program. Luiz et al. (2023) reiterate the importance of quarantines and education to curtail movement of infected material and countering activities that injure the trees. Fencing to protect these forests from grazing by feral animals can drastically reduce the amount of disease. Finally, scientists must overcome the factors there caused the almost complete lack of natural regeneration of ‘ōhi‘a in lower elevation forests. Most important are competition by invasive plants, predation by feral ungulates, and the presence of other diseases, e.g., Austropuccinia psidii.
Hawaii’s dryland forests are highly endangered: more than 90% of dry forests are already lost due to habitat destruction and the spread of invasive plant and animal species. Two tree species are the focus of species-specific programs aimed at restoring them to remaining dryland forests. However, support for both programs seems precarious and requires stable long-term funding; disease resistance programs often necessitate decades-long endeavors.
naio in bloom; photo by Forrest & Kim Starr via Creative Commons
1) naio (Myoporum sandwicense)
Naio grows on all of the main Hawaiian Islands at elevations ranging from sea level to 3000 m. While it occurs in the full range of forest types from dry to wet, naio is one of two tree species that dominate upland dry forests. The other species is mamane, Sophora chrysophylla. Naio is a key forage tree for two endangered honeycreepers, palila (Loxioides bailleui) and `akiapola`au (Hemignathus munroi). The tree is also an important host of many species of native yellow-face bees (Hylaeus spp). Finally, loss of a native tree species in priority watersheds might lead to invasions by non-native plants that consume more water or increase runoff.
The invasive non-native Myoporum thrips, Klambothrips myopori, was detected on Hawai‘i Island in December 2008 (L. Kaufman website). In 2018 the thrips was found also on Oahu (work plan). The Myoporum thrips feeds on and causes galls on plants’ terminal growth. This can eventually lead to death of the plant.
Aware of thrips-caused death of plants in the Myoporum genus in California, the Hawaii Department of Lands and Natural Resources Division of Forestry and Wildlife and the University of Hawai‘i began efforts to determine the insect’s distribution and infestation rates, as well as the overall health of naio populations on the Big Island. This initiative began in September 2010, nearly two years after the thrips’ detection. Scientists monitored nine protected natural habitats for four years. This monitoring program was supported by the USFS Forest Health Protection program. This program is described by Kaufman.
naio monitoring sites from L. Kaufman article
The monitoring program determined that by 2013, the thrips has spread across most of Hawi`i Island, on its own and aided by human movement of landscaping plants. More than 60% of trees being monitored had died. Infestation and dieback levels had both increased, especially at medium elevation sites. The authors feared that mortality at high elevations would increase in the future. They found no evidence that natural enemies are effective controlling naio thrips populations on Hawai`i Island.
Kaufman was skeptical that biological control would be effective. She suggested, instead, a breeding program, including hybridizing M. sandwicensis with non-Hawaiian Myoporum species that appear to be resistant to thrips. Kaufman also called for additional programs: active monitoring to prevent thrips from establishing on neighboring islands; and collection and storage of naio seeds.
Ten years later, in February 2024, DLNR Division of Forestry and Wildlife adopted a draft work plan for exploring possible resistance to the Myoporum thrips. Early steps include establishing a database to record data needed to track parent trees, associated propagules, and the results of tests. These data are crucial to keeping track of which trees show the most promise. Other actions will aim to hone methods and processes. Among practical questions to be answered are a) whether scientists can grow even-aged stands of naio seedlings; b) identifying the most efficient resistance screening techniques; and c) whether K. myopori thrips are naturally present in sufficient numbers to be used in tests, or – alternatively – whether they must be augmented. [Plan]
Meanwhile, scientists have begun collecting seed from unaffected or lightly affected naio in hotspots where mortality is high. They have focused on the dry and mesic forests of the western side of Hawai`i (“Big”) Island, where the largest number of naio populations still occur and are at high risk. Unfortunately, these “lingering” trees remain vulnerable to other threats, such as browsing by feral ungulates, competition with invasive plants, drought, and reduced fecundity & regeneration.
Hawai`i DLNR has secured initial funding from the Department of Defense’s REPI program to begin a pest resistance project and is seeking a partnership with University of Hawai`i to carry out tests “challenging” different naio families’ resistance to the thrips [R. Hauff pers. comm.]
wiliwili; photo by Forrest & Kim Starr
2) wiliwili (Erythrina sandwicensis)
Efforts to protect the wiliwili have focused on biological control. The introduced Erythrina gall wasp, Quadrastichus erythrinae (EGW) was detected on the islands in 2005. It immediately caused considerable damage to the native tree and cultivated nonnative coral trees.
A parasitic wasp, Eurytoma erythrinae, was approved for release in November 2008 – only 3 ½ years after EGW was detected on O‘ahu. The parasitic wasp quickly suppressed the gall wasp’s impacts to both wiliwili trees and non-native Erythrina. By 2024, managers are once again planting the tree in restoration projects.
However, both the gall wasp and a second insect pest – a bruchid, Specularius impressithorax – can cause loss of more than 75% of the seed crop. This damage means that the tree cannot regenerate. By 2019, Hawaiian authorities began seeking permission to release a second biocontrol gent, Aprostocitus nites.Unfortunately, the Hawai’i Department of Agriculture still has not approved the release permit despite five years having passed. Once they have this approval, the scientists will then need to ask USDA Animal and Plant Health Inspection Service (APHIS) for its approval [R. Hauff, pers. comm.]
SOURCES
www.RapidOhiaDeath.org
Dudley, N., R. James, R. Sniezko, P. Cannon, A. Yeh, T. Jones, & Michael Kaufmann. 2009? Operational Disease Screening Program for Resistance to Wilt in Acacia koa in Hawai`i. Hawai`i Forestry Association Newsletter August 29 2009
Dudley, N., T. Jones, K. Gerber, A.L. Ross-Davis, R.A. Sniezko, P. Cannon & J. Dobbs. 2020. Establishment of a Genetically Diverse, Disease-Resistant Acacia koa Seed Orchard in Kokee, Kauai: Early Growth, Form, & Survival. Forests 2020, 11, 1276; doi:10.3390/f11121276 www.mdpi.com/journal/forests
Friday, J. B., L. Keith, and F. Hughes. 2015. Rapid ʻŌhiʻa Death (Ceratocystis Wilt of ʻŌhiʻa). PD-107, College of Tropical Agriculture and Human Resources, University of Hawai‘i, Honolulu, HI. URL: https://www.ctahr.HI.edu/oc/freepubs/pdf/PD-107.pdf Accessed April 3, 2018.
Friday, J.B. 2018. Rapid ??hi?a Death Symposium -West Hawai`i (“West Side Symposium”) March 3rd 2018, https://vimeo.com/258704469 Accessed April 4, 2018 (see also full video archive at https://vimeo.com/user10051674)
Inman-Narahari, F., R. Hauff, S.S. Mann, I. Sprecher, & L. Hadway. Koa Action Plan: Management & research priorities for Acacia koa forestry in Hawai`i. State of Hawai`i Department of Land & Natural Resources Division of Forestry & Wildlife no date
Kaufman, L.V, J. Yalemar, M.G. Wright. In press. Classical biological control of the erythrina gall wasp, Quadrastichus erythrinae, in Hawaii: Conserving an endangered habitat. Biological Control. Vol. 142, March 2020
Loope, L. and A.M. LaRosa. 2008. ‘Ohi’a Rust (Eucalyptus Rust) (Puccinia psidii Winter) Risk Assessment for Hawai‘i.
Luiz, B.C. 2017. Understanding Ceratocystis. sp A: Growth, morphology, and host resistance. MS thesis, University of Hawai‘i at Hilo.
Luiz, B.C., C.P. Giardina, L.M. Keith, D.F. Jacobs, R.A. Sniezko, M.A. Hughes, J.B. Friday, P. Cannon, R. Hauff, K. Francisco, M.M. Chau, N. Dudley, A. Yeh, G. Asner, R.E. Martin, R. Perroy, B.J. Tucker, A. Evangelista, V. Fernandez, C. Martins-Keli’iho.omalu, K. Santos, R. Ohara. 2023. A framework for establishlishing a rapid ‘Ohi‘a death resistance program New Forests 54, 637–660. https://doi.org/10.1007/s11056-021-09896-5
Sniezko, R.A., N. Dudley, T. Jones, & P. Cannon. 2016. Koa wilt resistance & koa genetics – key to successful restoration & reforestation of koa (Acacia koa). Acacia koa in Hawai‘i: Facing the Future. Proceedings of the 2016 Symposium, Hilo, HI: www.TropHTIRC.org , www.ctahr.HI.edu/forestry
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
The number of introduced forest pathogens are increasing – creating a crisis that is recognized by more scientists. These experts say tree diseases are reshaping both native and planted forests around the globe. The diseases are threatening biodiversity, ecosystem services, provision of products, and related human wellbeing. Some suggest that bioinvasions might threaten forests as much as climate change, while also undermining forests’ role in carbon sequestration.
Unfortunately, I see little willingness within the plant health regulatory community to tackle improving programs to slow introductions. Even when the scientists documenting the damage work for the U.S. Department of Agriculture – usually the U.S. Forest Service — USDA policy-makers don’t act on their findings. [I tried to spur a conversation with USDA 2 years ago. So far, no response.]
counties where beech leaf disease has been detected
What the scientists say about these pests’ impacts
Andrew Gougherty (2023) – one of the researchers employed by the USDA Forest Service – says that emerging infectious tree diseases are reshaping forests around the globe. Furthermore, new diseases are likely to continue appearing in the future and threaten native and planted forests worldwide. [Full references are provided at the end of the blog.] Haoran Wu (2023/24) – a Master’s Degree student at Oxford University – agrees that arrival of previously unknown pathogens are likely to alter the structure and composition of forests worldwide. Weed, Ayers, and Hicke (2013) [academics] note that forest pests — native and introduced — are the dominant sources of disturbance to North American forests. They suggest that, globally, bioinvasions might be at least as important as climate change as threats to the sustainability of forest ecosystems. They are concerned that recurrent forest disturbances caused by pests might counteract carbon mitigation strategies.
Scientists have proclaimed these warnings for years. Five years ago, Fei et al. (2019) reported that the 15 most damaging pests introduced to the United States — cumulatively — had already caused tree mortality to exceed background levels by 5.53 teragrams of carbon per year. As these 15 pests spread and invasions intensify, they threaten 41.1% of the total live forest biomass in the 48 coterminous states. Poland et al. (2019) (again – written by USFS employees) document the damage to America’s forest ecosystems caused by the full range of invasive species, terrestrial and aquatic.
Fei et al. and Weed, Ayers, and Hicke (2013) also support the finding that old, large trees are the most important trees with regard to carbon storage. This understanding leads them to conclude that the most damaging non-native pests are the emerald ash borer, Dutch elm disease fungi, beech bark disease, and hemlock woolly adelgid. As I pointed out in earlier blogs, other large trees, e.g., American chestnut and several of the white pines, were virtually eliminated from much of their historical ranges by non-native pathogens decades ago. These same large, old, trees also maintain important aspects of biological diversity.
It is true that not all tree species are killed by any particular pest. Some tree genera or species decrease while others thrive, thus altering the species composition of the affected stands (Weed, Ayers, and Hicke). This mode of protection is being undermined by the proliferation of insects and pathogens that cumulatively attack ever more tree taxa. And while it is true that some of the carbon storage capacity lost to pest attack will be restored by compensatory growth in unaffected trees, this faster growth is delayed by as much as two or more decades after pest invasions begin (Fei et al.).
ash forest after EAB infestation; Photo by Nate Siegert, USFS
Still, despite the rapid rise of destructive tree pests and disease outbreaks, scientists cannot yet resolve critical aspects of pathogens’ ecological impacts or relationship to climate change. Gougherty notes that numerous tree diseases have been linked to climate change or are predicted to be impacted by future changes in the climate. However, various studies’ findings on the effects of changes in moisture and precipitation are contradictory. Wu reports that his study of ash decline in a forest in Oxfordshire found that climate change will have a very small positive impact on disease severity through increased pathogen virulence. Weed, Ayers, and Hicke go farther, making the general statement that despite scientists’ broad knowledge of climate effects on insect and pathogen demography, they still lack the capacity to predict pest outbreaks under climate change. As a result, responses intended to maintain ecosystem productivity under changing climates are plagued by uncertainty.
Clarifying how disease systems are likely to interact with predicted changes in specific characteristics of climate is important — because maintaining carbon storage levels is important. Quirion et al. (2021) estimate that, nation-wide, native and non-native pests have decreased carbon sequestration by live forest trees by at least 12.83 teragrams carbon per year. This equals approximately 9% of the contiguous states’ total annual forest carbon sequestration and is equivalent to the CO2 emissions from more than 10 million passenger vehicles driven for one year. Continuing introductions of new pests, along with worsening effects of native pests associated with climate change, could cause about 30% less carbon sequestration in living trees. These impacts — combined with more frequent and severe fires and other forest disturbances — are likely to negate any efforts to improve forests’ capacity for storing carbon.
Understanding pathogens’ interaction with their hosts is intrinsically complicated. There are multiple biological and environmental factors. What’s more, each taxon adapts individually to the several environmental factors. Wu says there is no general agreement on the relative importance of the various environmental factors. The fact that most forest diseases are not detected until years after their introduction also complicates efforts to understand factors affecting infection and colonization.
The fungal-caused ash decline in Europe is a particularly alarming example of the possible extent of such delays. According to Wu, when the disease was first detected – in Poland in 1992 – it had already been present perhaps 30 years, since the 1960s. Even then, the causal agent was not isolated until 2006 – or about 40 years after introduction. The disease had already spread through about half the European continent before plant health officials could even name the organism. The pathogen’s arrival in the United Kingdom was not detected until perhaps five years after its introduction – despite the country possessing some of the world’s premier forest pathologists who by then (2012) knew what they to look for.
Clearly, improving scientific understanding of forest pathogens will be difficult. In addition, effective policy depends on understanding the social and economic drivers of trade, development, and political decisions are primary drivers of the movement of pathogens. Wu calls for collaboration of ecologists, geneticists, earth scientists, and social scientists to understand the complexity of the host-pathogen-surrounding system. Bringing about this new way of working and obtaining needed resources will take time – time that forests cannot afford.
However, Earth’s forests are under severe threat now. Preventing their collapse depends on plant health officials integrating recognition of these difficulties into their policy formulation. It is time to be realistic: develop and implement policies that reflect the true level of threat and limits of current science.
Background: Rising Numbers of Introductions
Gougherty’s analysis of rising detections of emerging tree diseases found little evidence of saturation globally – in accord with the findings of Seebens et al. (2017) regarding all taxa. Relying on data for 24 tree genera, nearly all native to the Northern Hemisphere, Gougherty found that the number of new pests attacking these tree genera are doubling on average every 11.2 years. Disease accumulation is increasing rapidly in both regions where hosts are native and where they are introduced, but more rapidly in trees’ native ranges.This finding is consistent with most new diseases arise from introductions of pathogens to naïve hosts.
Gougherty says his estimates are almost certainly underestimates for a number of reasons. Countries differ in scientific resources and their scientists’ facility with English. Scientists are more likely to notice and report high-impact pathogens and those in high-visibility locations. Where national borders are closer, e.g., in Europe, a minor pest expansion can be reported as “new” in several countries. New pathogens in North America appear to occur more slowly, possibly because the United States and Canada are very large. He suggests that another possible factor is the U.S. (I would add Canada) have adopted pest-prevention regulations that might be more effective than those in place in other regions. (See my blogs and the Fading Forest reports linked to below for my view of these measures’ effectiveness.)
ash dieback in the UK
Wu notes that reports of tree pathogens in Europe began rising suddenly after the 1980s. He cites the findings by Santini et al. (2012) that not only were twice as many pathogens detected in the period after 1950 than in the previous 40 years, the region of origin also changed. During the earlier period, two-thirds of the introduced pathogens came from temperate North America. After 1950, about one-third of previously unknown disease agents were from temperate North America. Another one-third was from Asia. By 2012, more than half of plant infectious diseases were caused by introduction of previously unknown pathogens.
What is to be done?
Most emerging disease agents do not have the same dramatic effects as chestnut blight in North America, ash dieback in Europe, or Jarrah dieback in Australia. Nevertheless, as Gougherty notes, their continued emergence in naïve biomes increases the likelihood of especially damaging diseases emerging and changing forest community composition.
Gougherty calls for policies intended to address both the agents being introduced through trade, etc., and those that emerge from shifts in virulence or host range of native pathogens or changing environmental conditions. In his view, stronger phytosanitary programs are not sufficient.
Wu recommends enhanced monitoring of key patterns of biodiversity and ecosystem functioning, He says these studies should focus on the net outcome of complex interactions. Wu also calls for increasing understanding of key “spillover” effects – outcomes that cannot be currently assessed but might impact the predicted outcome. He lists several examples:
the effects of drought–disease interactions on tree health in southern Europe,
interaction between host density and pathogen virulence,
reproductive performance of trees experiencing disease,
effect of secondary infections,
potential for pathogens to gain increased virulence through hybridization.
potential for breeding resistant trees to create a population buffer for saving biological diversity. Wu says his study of ash decline in Oxfordshire demonstrates that maintaining a small proportion of resistant trees could help tree population recovery.
Quirion et al. provide separate recommendations with regard to native and introduced pests. To minimize damage from the former, they call for improved forest management – tailored to the target species and the environmental context. When confronting introduced pests, however, thinning is not effective. Instead, they recommend specific steps to minimize introductions via two principal pathways, wood packaging and imports of living plants. In addition, since even the most stringent prevention and enforcement will not eliminate all risk, Quirion et al. advocate increased funding for and research into improved strategies for inspection, early detection of new outbreaks, and strategic rapid response to newly detected incursions. Finally, to reduce impacts of established pests, they recommend providing increased and more stable funding for classical biocontrol, research into technologies such as sterile-insect release and gene drive, and host resistance breeding.
USDA HQ
Remember: reducing forest pest impacts can simultaneously serve several goals—carbon sequestration, biodiversity conservation, and perpetuating the myriad economic and societal benefits of forests. See Poland et al. and the recent IUCN report on threatened tree species.
SOURCES
Barrett, T.M. and G.C. Robertson, Editors. 2021. Disturbance and Sustainability in Forests of the Western United States. USDA Forest Service Pacific Northwest Research Station. General Technical Report PNW-GTR-992. March 2021
Clark, P.W. and A.W. D’Amato. 2021. Long-term development of transition hardwood and Pinus strobus – Quercus mixedwood forests with implications for future adaptation and mitigation potential. Forest Ecology and Management 501 (2021) 119654
Fei, S., R.S. Morin, C.M. Oswalt, and A.M. 2019. Biomass losses resulting from insect and disease invasions in United States forests. Proceedings of the National Academy of Sciences. www.pnas.org/cgi/doi/10.1073/pnas.1820601116
Gougherty AV (2023) Emerging tree diseases are accumulating rapidly in the native and non-native ranges of Holarctic trees. NeoBiota 87: 143–160. https://doi.org/10.3897/neobiota.87.103525
Lovett, G.M., C.D. Canham, M.A. Arthur, K.C. Weathers, and R.D. Fitzhugh. 2006. Forest Ecosystem Responses to Exotic Pests and Pathogens in Eastern North America. BioScience Vol. 56 No. 5 May 2006
Lovett, G.M., M. Weiss, A.M. Liebhold, T.P. Holmes, B. Leung, K.F. Lambert, D.A. Orwig, F.T. Campbell, J. Rosenthal, D.G. MCCullough, R. Wildova, M.P. Ayres, C.D. Canham, D.R. Foster, S.L. Ladeau, and T. Weldy. 2016. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecological Applications, 26(5), 2016, pp. 1437-1455
Poland, T.M., Patel-Weynand, T., Finch, D., Miniat, C. F., and Lopez, V. (Eds) (2019), Invasive Species in Forests and Grasslands of the United States: A Comprehensive Science Synthesis for the United States Forest Sector. Springer Verlag.
Quirion, B.R., G.M. Domke, B.F. Walters, G.M. Lovett, J.E. Fargione, L. Greenwood, K. Serbesoff-King, J.M. Randall, and S. Fei. 2021 Insect and Disease Disturbance Correlate With Reduced Carbon Sequestration in Forests of the Contiguous US. Front. For. Glob. Change 4:716582. [Volume 4 | Article 716582] doi: 10.3389/ffgc.2021.716582
Weed, A.S., M.P. Ayers, and J.A. Hicke. 2013. Consequences of climate change for biotic disturbances in North American forests. Ecological Monographs, 83(4), 2013, pp. 441–470
Wu, H. 2023/24. Modelling Tree Mortality Caused by Ash Dieback in a Changing World: A Complexity-based Approach MSc/MPhil Dissertation Submitted August 12, 2024. School of Geography and the Environment, Oxford University.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
I welcome new attention to the threats posed to tree species around world.
Last week, at the conclusion of Conference of the Parties (COP) to the Convention on Biodiversity (CBD), the International Union for the Conservation of nature (IUCN) released its most recent iteration of the Red List of Threatened Species. The headline was that 38% of the world’s trees are at risk of extinction.
This is the finding of a decade-long Global Tree Assessment. The assessment was led by Botanic Gardens Conservation International and IUCN’s Species Survival Commission Global Tree Specialist Group. Partners in the effort included Conservation International, NatureServe, Missouri Botanical Garden and Royal Botanic Gardens, Kew. The project was funded primarily by Fondation Franklinia. The foundation was formed in 2005 expressly to conserve threatened tree species! I regret that I had not heard about it before.
At least 16,425 of the 47,282 tree species assessed are at risk of extinction. Trees now account for over one quarter of species on the IUCN Red List, and the number of threatened trees is more than double the number of all threatened birds, mammals, reptiles and amphibians combined. Tree species are at risk of extinction in 192 countries around the world.
No surprise: the highest proportion of threatened trees is found on islands. Island trees are at particularly high risk due to deforestation for urban development, conversion to agriculture, invasive species, pests and diseases. Climate change is increasingly threatening trees, especially in the tropics, through sea-level rise and stronger, more frequent storms.
The COP was held in Cali, Columbia. This is fitting because South America is home to the greatest diversity of trees in the world. Twenty-five percent – 3,356 out of 13,668 assessed species are at risk of extinction. Forest clearance for crop farming and livestock ranching are the largest threats on the continent. Dr Eimear Nic Lughadha, Senior Research Leader in Conservation Assessment and Analysis at the Royal Botanic Gardens, Kew, said this percentage is sure to increase as many additional tree species are described for science.
IUCN spokespeople emphasized that the loss of trees is a major threat to thousands of other plants, fungi and animals. Cleo Cunningham, Head of Climate and Forests at Birdlife International pointed out that over two-thirds of globally threatened bird species are dependent on forests. Speakers also noted that people depend on trees; over 5,000 of the tree species on the Red List are used in construction, and over 2,000 species provide medicines, food and fuels.
Sam Ross, Sustainable Business Project Analyst at ZSL, noted that “Despite growing pressure to halt worldwide deforestation by 2030, … most of the world’s 100 most significant tropical timber and pulp companies have made limited progress in disclosing their zero deforestation and traceability commitments. We must all do more to safeguard these vital forest ecosystems, especially consumer goods manufacturers, financial institutions funding forestry, and agriculture companies.”
IUCN and the Red List Partners are launching a global social media campaign to raise awareness and funds to accelerate species assessments and reassessments. The campaign will culminate at the IUCN World Conservation Congress in Abu Dhabi, in October 2025.
Impacts from Pathogens Continue to Increase
Meanwhile, in North America and elsewhere, infections by tree-killing pathogens are spreading and intensifying.
In California, P. ramorum the statewide rate of tree infection in 2024 doubled from 2023. Expansions were most obvious in Mendocino and Del Norte counties. Worse, California has now detected a third strain of P. ramorum in its forests. The NA2 strain was first detected in Del Norte County in 2020. Now it has been found in five sites closer to the “core” of the infestation closer to San Francisco Bay. Dr. Matteo Garbelotto believes the strain – formerly known only in nurseries – had been present for some years. It appears to be more aggressive than the strain long present in forests – NA1 – and might be favored by warmer temperatures. [The EU1 strain was detected in Del Norte County in 2021.]
Oregon has been wrestling with the EU1 strain since 2015; the NA2 strain since 2021. Beginning in late 2022, authorities have discovered multiple disease outbreaks between the Rogue River and Port Orford (farther north than the area previously known to be infected). Many of these new outbreaks are the EU1 lineage. The state is struggling to carry out eradication treatments using funds from state legislative appropriations, support from USDA Forest Service and USDI Bureau of Land Management, USDA Agriculture Research Service, and direct Congressional appropriations. The last resulted from assertive lobbying!
The Government Accountability Office is studying interactions between climate change and agricultural pests; sudden oak death is one of four focal pests. The report is expected to be released in 2025.
[Most of this information is from the California Oak Mortality Task Force (COMTF) webinar on 29 October, 2024. Recording available here.]
limber pine in Rocky Mountain National Park; photo by F.T. Campbell
Limber pine (Pinus flexilis) is heavily infected by blister rust in Alberta; in its U.S. range
range of limber pine
the disease is increasing. Scientists had been cheered by the presence of major gene resistance (MGR) in limber pine to the rust. However, a strain of blister rust in Alberta has been determined to be virulent despite this gene (Liu et al. 2024). Scientists might have to launch a breeding program to try to enhance quantitative disease resistance (QDR) in the species. Unfortunately, the frequency and level of partial resistance in limber pine has been very low in trees tested so far. Scientists now must test more limber pines to see whether some have higher levels of QDR.
Southwestern white pine (Pinus strobiformis) presents the same problem; the MGR gene might even be the same gene. Some some populations of SWWP have higher partial or quantitative disease resistance.
beech leaf disease in southern Fairfax County, Virginia; photo by F.T. Campbell (apologies for the quality)
BLD continues to be detected in new sites. According to Matthew Borden of Bartlett Tree Research Laboratories, since 2021, BLD has been detected in five counties in Virginia:
Prince William County — Prince William Forest Park;
Fairfax County: southern Fairfax County on the border with Prince William County (Fountainhead Park, Hemlock Overlook Park, and Meadowood Special Recreation Area), somewhat farther north (Burke Lake Park), and northern edge (Great Falls);
Loudoun County;
Stafford County – just outside the city of Fredricksburg and along the Spotsylvania river
New Kent County in Wahrani Natural Preserve
Several of these outbreaks – e.g., southern Fairfax County, Stafford County, and Loudoun County – are 20 miles or more away from other known outbreaks. Virginia Department of Agriculture staff are monitoring the disease. All these sites are near water – although the Potomac River in Loudoun County is above the fall line so narrower than at the other sites.
SOURCE
Liu, J-J., R.A. Sniezko, S. Houston, G. Alger, J. Krakowski, A.W. Schoettle, R. Sissons, A. Zamany, H. Williams, B. Rancourt, A. Kegley. 2024. A New Threat to Limber Pine (Pinus flexilis) Restoration in Alberta and Beyond: First Documentation of a Cronartium ribicola race (vcr4) Virulent to Cr4-Controlled Major Gene Resistance. Phytopathology. Published Online:25 Sep 2024 https://doi.org/10.1094/PHYTO-04-24-0129-R
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
I learned at the beginning of August that Canadian scientists have discovered a new pathogen causing wilt disease on American elms (Ulmus americana). The pathogen is Plenodomus tracheiphilus, which is known primarily for causing serious disease in citrus.
P. tracheiphilus is described as common on Alberta’s elm trees, especially in the Edmonton area. It was found on 116 of 200 trees which were sampled – see map. The wilting had previously been blamed on Dothiorella ulmi. I have been unable to find a source for the geographic origin of Dothiorella ulmi; perhaps it is native to North America. It is reported to be present at least from Alberta to Texas. (Presumably if Plenodomus tracheiphilus were in Texas it would have caused obvious symptoms on that state’s citrus crops.)
poster prepared by Alberta Plant Health Lab, Alberta Agriculture & Irrigation, and Society to Prevent Dutch Elm Disease
I am unaware of any North American forest pathologists studying whether this pathogen is also established in the United States, or its possible effects. The discovery in Alberta is the first time this organisms has been associated with disease on elms; I have asked European and North American forest pathologists whether they are looking into possible disease on any of the European or North American elm species. So far, no one reports that s/he has been.
In the meantime, the California Department of Food and Agriculture has begun the process of assigning Plenodomus tracheiphilus the highest pest risk designation for the state. CDFA is worried primarily about damage to the state’s $2.2 billion citrus industry. CDFA is seeking comments on its proposed action; go here .
CDFA points out that despite awareness of the disease on economically important citrus since at least 1900 and efforts by phytosanitary agencies, it has spread to most citrus-growing countries around the Mediterranean and Black seas and parts of the Middle East. The primary mode of spread is movement of infected plant material, e.g., rootstocks, grafted plants, scions, budwood, and even fruit peduncles and leaves. Transmission is possible from latently infected, asymptomatic material. Once established at a site, the conidia produced on diseased plant parts can be spread over relatively short distances by rain-splash, overhead irrigation, water surface flow, or wind-driven rain. Transport by birds and insects is also suspected. The pathogen can survive on pruned material or in soil containing infected plant debris for up to four month.
The report from Canada does not speculate on how a disease associated with plants in a Mediterranean climate was transported to Alberta, which has a cold continental climate. Nor is there any information on the possible presence of the disease on elms in warmer parts of Canada.
U.S. elms appear to be at high risk because phytosanitary restrictions leave dangerous gaps.
First, under the Not Authorized for Importation Pending Pest Risk assessment (NAPPRA) program, USDA APHIS has prohibited importation of plants in the Ulmus genus from all countries except Canada. Second, importation of cut greenery is allowed from all countries – and the CDFA analysis indicates that the pathogen can be transported on leaves. Third, it appears to me that it is probable that this pathogen survives on plants in additional taxa.
See this profile for a description of other threats to North American elms.
Yang, Y., H. Fu, K. Zahr, S. Xue, J. Calpas, K. Demilliano, et al. 2024. Plenodomus tracheiphilus, but not Dothiorella ulmi, causes wilt disease on elm trees in Alberta, Canada. European Journal of Plant Pathology 169(2):409-420. Last accessed August 1, 2024, from https://link.springer.com/article/10.1007/s10658-024-02836-x
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
Research scientists in the USFS Northern Region (Region 9) – Maine to Minnesota, south to West Virginia and Missouri – continue to be concerned about regeneration patterns of the forest and the future productivity of northern hardwood forests.
The most recent study of which I am aware is that by Stern et al. (2023) [full citation at the end of this blog]. They sought to determine how four species often dominant in the Northeast (or at least in New England) might cope with climate change. Those four species are red maple (Acer rubrum), sugar maple (Acer saccharum), American beech (Fagus grandifolia), and yellow birch (Betula alleghaniensis). The study involved considerable effort: they examined tree ring data from 690 dominant and co-dominant trees on 45 plots at varying elevations across Vermont. The tree ring data allowed them to analyze each species’ response to several stressors over the 70-year period of 1945 to 2014.
In large part their findings agreed with those of studies carried out earlier, or at other locations. As expected, all four species grew robustly during the early decades, then plateaued – indicative of a maturing forest. All species responded positively to summer and winter moisture and negatively to higher summer temperatures. Stern et al. described the importance of moisture availability in non-growing seasons – i.e., winter – as more notable.
snow in Vermont; Putnypix via Flickr
The American Northeast and adjacent areas in Canada have already experienced an unprecedented increase of precipitation over the last several decades. This pattern is expected to continue or even increase under climate change projections. However, Stern et al. say this development is not as promising for tree growth as it first appears. The first caveat is that winter snow will increasingly be replaced by rain. The authors discuss the importance of the insulation of trees’ roots provided by snow cover. They suggest that this insulation might be particularly necessary for sugar maple.
The second caveat is that precipitation is not expected to increase in the summer; it might even decrease. Their data indicate that summer rainfall – during both the current and preceding years – has a significant impact on tree growth rates.
Stern et al. also found that the rapid rise in winter minimum temperatures was associated with slower growth by sugar maple, beech, and yellow birch, as well as red maple at lower elevations. Still, temperature had less influence than moisture metrics.
Stern et al. discuss specific responses of each species to changes in temperatures, moisture availability, and pollutant deposition. Of course, pollutant levels are decreasing in New England due to implementation of provisions of the Clean Air Act of 1990.
They conclude that red maple will probably continue to outcompete the other species.
In their paper, Stern et al. fill in some missing pieces about forests’ adaptation to the changing climate. I am disappointed, however, that these authors did not discuss the role of biotic stressors, i.e., “pests”.
They do report that growth rates of American beech increased in recent years despite the prevalence of beech bark disease. They note that others’ studies have also found an increase in radial growth for mature beech trees in neighboring New Hampshire, where beech bark disease is also rampant.
For more specific information on pests, we can turn to Ducey at al. – also published in 2023. These authors expected American beech to dominate the Bartlett Experimental Forest (in New Hampshire) despite two considerations that we might expect to suppress this growth. First, 70-90% of beech trees were diseased by 1950. Second, managers have made considerable efforts to suppress beech.
Stern et al. say specifically that their study design did not allow analysis of the impact of beech bark disease. I wonder at that decision since American beech is one of four species studied. More understandable, perhaps, is the absence of any mention of beech leaf disease. In 2014, the cutoff date for their growth analysis, beech leaf disease was known only in northeastern Ohio and perhaps a few counties in far western New York and Pennsylvania. Still, by the date of publication of their study, beech leaf disease was recognized as a serious disease established in southern New England.
counties where beech leaf disease has been confirmed
Eastern hemlock (Tsuga canadensis) and northern red oak (Quercus rubra) are described as common co-occurring dominant species in the plots analyzed by Stern et al. Although hemlock woolly adelgid has been killing trees in southern Vermont for years, Stern et al. did not discuss the possible impact of that pest on the forest’s regeneration trajectory. Nor did they assess the possible effects of oak wilt, which admittedly is farther away (in New York (& here) and in Ontario, Canada, west of Lake Erie).
In contrast, Ducey at al. (2023) did discuss link to blog 344 the probable impact of several non-native insects and diseases. In addition to beech bark disease, they addressed hemlock woolly adelgid, emerald ash borer, and beech leaf disease.
Non-native insects and pathogens are of increasing importance in our forests. To them, we can add overbrowsing by deer, proliferation of non-native plants, and spread of non-native earthworms. There is every reason to think the situation will only become more complex. I hope forest researchers will make a creative leap – incorporate the full range of factors affecting the future of US forests.
I understand that such a more integrated, holistic analysis might be beyond any individual scientist’s expertise or time, funding, and constraints of data availability and analysis. I hope, though, that teams of collaborators will compile an overview based on combining their research approaches. Such an overview would include human management actions, climate variables, established and looming pest infestations, etc. I hope, too, that these experts will extrapolate from their individual, site-specific findings to project region-wide effects.
Some studies have taken a more integrative approach. Reed, Bronson, et al. (2022) studied interactions of earthworm biomass and density with deer. Spicer et al. (2023) examined interactions of deer browsing and various vegetation management actions. Hoven et al. (2022) considered interactions of non-native shrubs, tree basal area, and forest moisture regimes.
Stern, R.L., P.G. Schaberg, S.A. Rayback, C.F. Hansen, P.F. Murakami, G.J. Hawley. 2023. Growth trends and environmental drivers of major tree species of the northern hardwood forest of eastern North America. J. For. Res. (2023) 34:37–50 https://doi.org/10.1007/s11676-022-01553-7
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
Oregon ash dominate wetlands of Ankeny NWR; photo by Wyatt Williams, Oregon Department of Forestry
One of these insects is the emerald ash borer (EAB). We easterners have “been there & done that”. However, programs aimed at conserving wetlands and riparian areas of the Western states – and the associated species — are at least as vulnerable to loss of ash. Worse, other tree taxa, specifically oaks, and the open woodlands they inhabit — are also under threat. The ecological tragedies continue to affect ever more forests.
|Emerald Ash Borer in Oregon and British Columbia
The emerald ash borer (EAB; Agrilus planipennis) was detected in Oregon in June 2022. Officials had been expecting an introduction and had begun preparations. Unsurprisingly, the infestation is more widespread than known at first: detections in two new locations, fairly close to the original in Forest Grove, mean the infested area now occupies three neighboring counties — Washington, Yamhill, and Marion counties.
Oregon officials are trying to slow spread of EAB by removing infested trees. Surveys in Washington County had identified 190 infested ash trees; 80 were removed in April 2024. They treated healthy ash trees in Washington County with injections of the systemic insecticide emamectin benzoate. The effort was already a daunting task: the survey had disclosed 6,500 ash trees in the vicinity. The city of Portland – only 25 miles away – has 94,000 ash trees (Profita 2024).
In May, 2024 EAB was detected in the city of Vancouver in British Columbia. This detection in the sixth Canadian province adds to the threat to the ecosystems of the region. The Canadian Food Inspection Agency (CFIA) now regulates the movement of all ash material such as logs, branches, and woodchips, and all species of firewood, from the affected sites.
The CFIA is also conducting surveillance activities to determine where EAB might be present, and is collaborating with the City of Vancouver, the Vancouver Board of Parks and Recreation, the Province of British Columbia, and other stakeholders to respond to the detections and slow the spread of this pest.
Importance of Oregon ash (Fraxinus latifolia)
The Oregon ash is the only ash species native to the Pacific Northwest. Its range stretches from southern British Columbia to so California, where it has hybridized with velvet ash (F. velutina). It is highly susceptible to EAB attack; there is a high probability that Oregon ash could be rendered functionally extinct (Maze, Bond and Mattsson 2024). This vulnerability prompted the International Union for Conservation of Nature (IUCN) to classify Oregon ash as “near threatened” as long ago as 2017 (Melton et al. 2024).
Oregon ash typically grows in moist, bottomland habitats. There it is a late-successional climax species. In Oregon’s Willamette Valley and Washington’s Puget Trough, the tree improves streams’ water quality by providing shade, bank stabilization, and filtration of pollutants and excess nutrients. Maintaining these ecological services is particularly important because these streams are crucial to salmonids (salmon and trout) and other native aquatic species (Maze, Bond and Mattsson 2024).
So it is not surprising that one component of Oregonians’ pre-detection preparations was an analysis of the likely impact of widespread ash mortality on populations of salmon, trout, and other aquatic species. I summarize the key findings of Maze, Bond and Mattsson here.
According to this study, salmonids and other cold-water aquatic species suffer population declines and health effects when stream water temperatures are too warm. A critical factor in maintaining stream temperatures is shade – usually created by trees. In the Pacific Northwest many streams’ temperatures already exceed levels needed to protect sensitive aquatic species. A key driver of increased stream temperatures – at least in the Willamette Basin – is clearing of forests to allow agriculture.
Decreasing streams’ temperatures is not only a good thing to do; it is legally required by the Endangered Species Act because several salmon and steelhead trout species are listed. In one response, the Oregon Department of Environmental Quality recommends restoration and protection of riparian vegetation as the primary methods for increasing stream shading and mitigating increased stream temperatures in the lower Willamette Basin.
The forests shading many low-elevation forested wetlands and tributaries of the Willamette and lower Columbia rivers are often composed exclusively of Oregon ash. Loss of these trees’ shade will affect not just the immediate streams but also increase the temperature of mainstem waterways downstream.
Oregon ash – EAB detection site; photo by Wyatt Williams, Oregon Department of Forestry
Replacements for Oregon Ash?
The magnitude of the ecological impacts of ash mortality in the many forested wetlands in the Willamette Valley will largely be determined by what plant associations establish after the ash die. Oregon ash is uniquely able to tolerate soils inundated for extended periods. No native tree species can fill the void when the ash die. Oregon white oak (Quercus garryana), black cottonwood (Populus trichocarpa), and the alders (Alnus rubra and A. rhombifolia), are shade intolerant and unlikely to persist in later seral stages in some settings.
If non-native species fill the gaps, they will provide inferior levels of ecosystem services – I would think particularly regarding wildlife habitat and invertebrate forage. Maze, Bond and Mattsson expect loss of ash to trigger significant physical and chemical changes. These will directly impact water quality and alter native plant and animal communities’ composition and successional trajectories.
The authors cite expectations of scientists studying loss of black ash (F. nigra) from upper Midwestern wetlands. There, research indicates loss of ash from these systems is likely to result in higher water tables and a conversion from forested to graminoid- or shrub-dominated systems. Significant changes follow: to food webs, to habitat structure, and, potentially, to nitrogen cycling.
Maze, Bond and Mattsson expect similar impacts in Willamette Valley wetlands and floodplains, especially those with the longest inundation periods and highest water tables. That is, there will probably be a broad disruption of successional dynamics and, at many sites, a conversion to open, shrub-dominated systems or to wetlands invaded by exotic reed canary grass (Phalaris arundinacea), with occasional sedge-dominated (Carex obnupta) wetlands. They think this change is especially likely under canopies composed of Oregon white oak (see below). The authors admit some uncertainty regarding the trajectories of succession because 90 years of water-control projects has almost eliminated the possibility of high-intensity floods.
Steelhead trout
Oregon Ash and Salmonids
Maze, Bond and Mattsson point out that all salmonids that spawn in the Willamette basin and the nearly 250,000 square mile extent of the Columbia basin upstream of Portland pass through the two wooded waterways in the Portland area that they studied. Applying a model to simulate disappearance of ash from these forests, the authors found that the reduced shade would raise the “solar load” on one waterway, which is wide and slow-moving, by 1.8%. On the second, much narrower, creek (mean channel width of 7 m), solar load was increased by of 23.7%.
Maze, Bond and Mattsson argue that even small changes can be important. Both waterbodies already regularly exceed Oregon’s target water temperature throughout the summer. Any increase in solar loading and water temperatures will have implications for the fish – and for entities seeking to comply with Endangered Species Act requirements. These include federal, state, and local governments, as well as private persons.
The Willamette and lower Columbia Rivers, and their tributaries, traverse a range of elevations. Ash trees comprise a larger proportion of the trees in the low elevation riparian and wetland forests. Consequently, Maze, Bond and Mattsson expect that EAB-induced loss of Oregon ash will have significant impacts on these rivers’ water quality and aquatic habitats. The higher water temperatures will affect aquatic organisms at multiple trophic levels.
They conclude that the EAB invasion West of the Cascade Mountain range constitutes an example of the worst-case forest pest scenario: the loss of a dominant and largely functionally irreplaceable tree species that provides critical habitat for both ESA-listed and other species, along with degradation of ecosystem services that protect water quality.
Breeding Oregon Ash … Challenges to be Overcome
According to Melton et al. (2024), Oregon ash does not begin to reproduce until it is 30 years old. Such an extended reproductive cycle could complicate breeding efforts unless scientists are able to accelerate flowering or use grafting techniques to speed up reproduction – as suggested by Richard Sniezko, USFS expert on tree breeding.
Melton et al. (2024) note that the IUCN has recently highlighted the importance of maintaining a species’ genetic variation in order to maintain its evolutionary potential. Consequently, they examined genetic variation in Oregon ash in order to identify the species’ ability to adjust to both the EAB threat and climate change. The authors sequenced the genomes of 1,083 individual ash trees from 61 populations. These spanned the species’ range from Vancouver Island to southern California. The genetic analysis detected four genetic clusters:
British Columbia;
Washington to central Oregon – including the Columbia River and its principal tributaries;
Southwest Oregon and Northwest California — the Klamath-Siskiyou ecoregion; and
all other California populations.
Connectivity between populations (that is, the potential corridors of movement for pollen and seeds and hence, genetic flow) was greatest in the riparian areas of the Columbia River and its tributaries in the center to the species’ range. Despite this evidence of connectivity, nucleotide diversity and effective population size were low across all populations. This suggests that the patchy distribution of Oregon ash populations might reduce its long-term evolutionary potential. As average temperatures rise, the regional populations will become more distinct genetically. The species’ ability to adjust to future climate projections is most constrained in populations on Vancouver Island and in smaller river valleys at the eastern and western edges of the range. Populations in southern California might be “pre-adapted” to warmer temperatures.
The resulting lower effective population size might exacerbate risks associated with EAB. The authors warned that although seeds from more than 350 maternal parent trees have been preserved since 2019, these collections do not cover the full genomic variation across Oregon ash’s range. Some genomic variation that represents adaptive variation critical to the species’ long-term evolution might be missing. They advocate using the genetic data from their study to identify regions where additional collections of germplasm are needed for both progeny trials and for long-term conservation.
Oregon white oak with symptoms of Mediterranean oak borer infestation; photo by Christine Buhl, Oregon Department of Forestry
Oregon White Oak (Quercus garryana) and the Mediterranean Oak Borer
The U.S. Department of Interior has been working with regional partners for 10 years to protect oak and prairie habitat for five ESA-listed species, two candidate species, and numerous other plant and animal species of concern. In August 2025 the Department announced creation of the Willamette Valley Conservation Area. It becomes part of the Willamette Valley National Wildlife Refuge Complex. These units are managed predominantly to maintain winter habitat for dusky geese (a separate population of Canada geese). Other units in the Complex are William L. Finley National Wildlife Refuge, Ankeny National Wildlife Refuge, and Baskett Slough National Wildlife Refuge.
These goals too face threats from non-native forest pests. First, the forested swamps of Ankeny NWR are composed nearly 100% of ash.
Second, Oregon white oak now confronts its own non-native pest – the Mediterranean oak borer (Xyleborus monographus). This Eurasian ambrosia beetle has been introduced to the northern end of the Willamette Valley (near Troutville, Oregon). It is likely that infestations are more widespread. Authorities are surveying areas near Salem. A separate introduction has become established in California, north of San Francisco Bay plus in Sacramento County in the Central Valley. Oregon white oak is vulnerable to at least one of the fungi vectored by this borer – Raffaelea montety. https://www.dontmovefirewood.org/pest_pathogen/mediterranean-oak-borer/
SOURCES
Maze, D., J. Bond and M. Mattsson. 2024. Modelling impacts to water quality in salmonid-bearing waterways following the introduction of emerald ash borer in the Pacific Northwest, USA. Biol Invasions (2024) 26:2691–2705 https://doi.org/10.1007/s10530-024-03340-3
Melton, A.E., T.M. Faske, R.A. Sniezko, T. Thibault, W. Williams, T. Parchman, and J.A. Hamilton. 2024. Genomics-driven monitoring of Fraxinus latifolia (Oregon Ash) for conservation and emerald ash borer resistance breeding. https://link.springer.com/article/10.1007/s10530-024-03340-3
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
beech leaf disease symptoms; photo by Matthew Borden via Flickr
Beech leaf disease (BLD) came to attention in 2012 near Cleveland. It has since spread to the Atlantic – Maine to New Jersey and northern Delaware; south into Virginia; north in Ontario; and west to eastern Michigan.
Scientists have scrambled to understand the disease – how it hijacks the tree’s metabolism; & here its impacts on seedlings, saplings, and mature trees; how it spreads, locations at greatest risk.
Now Bartlett Tree Research Laboratories – the research arm of Bartlett Tree Experts – has announced development of Integrated Pest Management (IPM) strategies to treat individual trees – sadly not yet beech in the forest. The project is led by Dr. Andrew Loyd and Dr. Matthew Borden.
Seeing the disease’s impacts on a tree species with aesthetic and ecological values not easily replaced, and its rapid spread, scientists at Bartlett Tree Research Laboratories began testing fungicides and nematicides registered under the Federal Insecticide, Fungicide and Rodenticide Act (FIFRA) by the U.S. Environmental Protection Agency (EPA) to see whether they might be effective against the causal nematode Litylenchus crenatae ssp mccannii.
As Drs. Loyd and Borden note, managing BLD presents numerous challenges:
1. The disease was discovered recently, so there were many unknowns, including how it spreads and the causal organism’s novel life cycle.
2. The damage occurs in leaf buds during winter dormancy. There has been little previous research on such a system. It is difficult for chemicals to reach the tissues.
3. Mature trees are large, so reaching the vulnerable leaves in the canopy is difficult.
4. Treatment efficacy cannot be evaluated until nearly a year after application.
5. Few chemicals are registered for use against foliar nematodes or for trees in forest, nursery, or landscape settings.
6. Obtaining funding is difficult because protecting beech is a low priority among many of the usual sources.
Fortunately, the leadership at Bartlett – the company’s research department, the New England Division leadership, and especially Robert A. Bartlett, Jr. (head of the family-owned company) – saw the importance of protecting beech and have supported this research. The USDA Forest Service has also funded some of studies exploring soil drenches. Cameron McIntire reports that these studies do not yet have results.
Furthermore, Bartlett has chosen to make the science easily available to all interested parties. Three posters explaining experiments to date are available at ResearchGate. They have also published a study on the early tests of Fuopyram as a foliar spray. It is open-access. Additional publications presenting data on experiments with both spray (Fluopyram) and injection (Thiabendazole/Arbotect) are in preparation.
I summarize briefly here their findings as of August 2024.
In all the trials, the scientists judged efficacy of treatments by counting the number of viable nematodes in leaves, viable nematodes in overwintering buds, and BLD symptom severity at appropriate times before and after treatment (spray or injection).
Tests of foliar sprays on small to medium sized trees
The first tests of foliar applications that resulted in BLD suppression were carried out in Ohio starting in 2021, then expanded to other field sites in Ohio and several states in New England in 2022 and 2023 seasons. In early trials, trees were sprayed four times starting in mid to late July at 21-day intervals. The scientists say that recent trials focus on application timing and rate. They hope that optimizing these factors will help generate new recommendations that are more sustainable while maintaining efficacy.
At the annual meeting of the American Phytopathological Society in July 2023, Bartlett announced that Fluopyram is an effective management tool to combat BLD – on smaller trees that can be treated using foliar application. There are several EPA-registered products, though only one, Broadform, has been so far been granted a section 2(EE) recommendation “For Control of Beech Leaf Disease on Beech Trees.”
Treatments are less effective in situations where the inoculum load is very high (for example, a very dense stand of infected trees); or where mature, untreated canopies hang over treated understory beech.
They suggest that managers focus treatments on high-value specimen beech, collection preservation, and potentially uncrowded mixed natural stands.
Treatments should be made by certified pesticide applicators who are familiar with the disease and treatment specifications. For the injection treatment, technical training and specialized equipment is needed. Bartlett arborists and plant health care specialists in locations affected by BLD have all been trained to perform the treatments, and some other arborists are doing BLD treatments as well using the same products.
Soil drench
Matt Borden said that they tested drenches with three different chemicals. The approach did not reduce nemtatode numbers sufficiently. However, as noted above, the Forest Service is funding additional tests exploring possible combinations of drenches with other actions, such as thinning. Discovering management options across a range of application methods (e.g., foliar, injection, drench) and modes of action is vital for a disease that covers such a broad range of locations and tree sizes and forms.
a macroinjection demonstration; photo by Matthew Borden via Flickr
Injections
Scientists injected Thiabendazole (TBZ) into beech on private land in three locations in Ohio and New Jersey. They tested two application rates and three application timings. They have two years of follow-up data for one site, one year for the others.
Key findings:
nematode numbers in buds in late winter consistently reflected foliar symptoms when the leaves opened.
Injections made before mid-July provided the greatest reduction in nemtatode numbers and best canopy improvement. Trees injected late in the season (30 August), after the nematode has begun dispersing from leaves to buds, exhibited some BLD symptoms the next year, but suffered less canopy dieback than controls.
Margery Daughtrey of Cornell said during a discussion of these finding that the trees’ persistence suggests that trees can tolerate some level of symptoms. Among other things, it might be possible to treat the trees less frequently than annually.
TBZ appears to provide at least two seasons of nematode suppression
Bartlett continues to monitor these trees to see how long the injected chemical suppresses nematode numbers and how long the tree remains healthy. They are also establishing new field sites to further optimize rate and timing.
TBZ – in a product called Arbotect 20-S – has been used to manage Dutch elm disease and sycamore anthracnose since the 1970s. However, it is also a well-known nematicide, previously used as an anti-parasitic drug in human and veterinary medicine. Once injected, TBZ protects the tree for more than one season. The injection technology (MACRO-Injection) has also been used for decades. It infuses the chemical directly into the tree’s vascular system; it does not rely on root uptake. Matt says injection does require take technical skill and the right equipment. To minimize the risk of the wound cracking and weeping, the injection should be done low on the side of the root flare, not on top.
While Arbotect 20-S has been registered for use in 48 states for many years, new labeling is required for its use in beech trees and against BLD. Special Local Needs labels, 24(C)s, have been granted by eight states – Connecticut, Massachusetts, Maine, New Jersey, New York, Pennsylvania, and Virginia. Registration in a ninth – Maryland – is in progress and Bartlett scientists are prepared to apply for several more. The problem is that only a limited number of these “special needs” labels may be issued, and BLD has expanded so far, and so rapidly, that it is already infesting beech in more states than may be covered by 24(C)s. Furthermore, 24(C) labels expire if not renewed. Most current 24(C)s will be active through 2028 – not ideal for a disease that will likely be with us long into the future. The product manufacturer (Syngenta) and distributor (Rainbow Ecoscience) are drafting a change to the main Arbotect 20-S label to add beech and the new nematode pest, but warn that EPA review and approval of amendments can take a very long time. Until then, we must resort to limited special local needs labels, and some states will miss out.
contrasting canopy transparency in beech treated with TBZ v. untreated controls; photo by Matthew Borden
One of the key scientists who developed these treatments for Dutch elm disease, R. Jay Stipes, professor emeritus at Virginia Tech, is quoted by Bartlett rejoicing that his work might help protect another tree species.
Matt believes the treatments will be effective if applied every 2-3 years. This approach would also spread out the cost – which will depend on the arborist but Dave Anderson of Rainbow Ecoscience estimated to be about $25 / inch of dbh.
It is always best to obtain an accurate diagnosis before treatment. The next step is talking through your options with a certified arborist or tree disease specialist. The “good” thing about BLD is that it is a progressive disease and will not kill a tree in a single year. Therefore, waiting until you know the disease is present or active locally is generally recommended.
Tree injection is better than foliar application where the latter is impractical (e.g., the tree is tall) or to reduce runoff, particularly near streams. Bartlett recommends treating any beech larger than 10 cm dbh by injection; smaller trees by foliar spray.
Treated trees should be sound, without serious decay, girdling roots, or other conditions that curtail uptake. Based on research results to date, they recommend treating the tree before mid-July. Bartlett is testing the results of injecting the shortly after full leaf expansion – early to mid-June. Bartlett scientists are testing several application rates to determine how long a single injection will suppress BLD. So far they have had good results from both low and moderate label rates (0.4-1.6 fl oz/inch DBH).
All the technical information re: research into treatments and recommendations for applying either the foliar or injection treatments has been provided by Dr. Matthew Borden of Bartlett Tree Research Laboratories. He can be reached at
Dr. Borden says he is immensely grateful for the support that allows him and Dr. Loyd to travel widely to establish the BLD research sites and spend weeks collecting data each year with their team. Company founder Francis A. Bartlett established the Bartlett Tree Research Laboratories as a separate entity within the company, where capital is reinvested directly into stable, long-term support of scientific tree research and preservation. The model is well-suited to provide the flexibility and freedom needed to rapidly respond to emerging invasive species issues.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/
The National Plant Board’s members are the lead plant health officials of the states and territories. Many federal officials also attend – from APHIS and DHS Bureau of Customs and Border Protection. Representatives of other North American phytosanitary entities participate – i.e., Canada, Mexico, and the North American Plant Protection Organization (NAPPO). Some stakeholder groups participate, especially the nursery industry. I have attended these meetings for over a decade because they provide an overview of pest issues and programs plus an unparalleled opportunity to network. The Nature Conservancy’s Leigh Greenwood also attends. We are the only representatives of the species conservation community to attend – others are missing great opportunities.
Here, I’ve listed 10 items that are among the most important the group discussed.
1) The funding situation for APHIS is worse than I realized
APHIS Administrator Mike Watson and Deputy Administrator (for plants) Mark Davidson both spoke about the need to cut programs to stay within the limits set by congressional appropriations. Funding for APHIS, as a whole, was cut only 1% for the current year (Fiscal Year 2024), cost-of-living salary increases mean less money for programs. (I believe Dr. Watson said $41 million less for FY24). If FY25 funding is the same, Congressionally mandated additional payraises will mean an another $20 million decrease in program funding.
Dr. Davidson said that the plant programs (Plant Protection and Quarantine) had been cut by 5% in FY24. However, Congress did not finalize the funding levels until about half-way through the fiscal year – so staying within the limits required even more severe cuts to programs in the remainder of FY24. To stay within these limits, APHIS cut several programs, among them a $3.6 million cut from the “tree and wood pest” program. This meant loss of funds to manage the polyphagous and Kuroshio shot hole borers, smaller cuts for programs managing Asian longhorned beetle and emerald ash borer, and perhaps the Asian flighted spongy moth. They anticipate additional cuts in these programs in FY25. The one bright light is the Section 7721 Plant Pest & Disease Management & Disaster Prevention Program. It provides steady funding for a range of plant health programs. The House version of the still-pending Farm Bill calls for increasing funding for this program by $15 million each year.
Nearly 100% ash trees in Oregon wetland — exposed to spreading EAB. Photo by Wyatt Williams, Oregon Department of Forestry
Remember this when I ask you to lobby for appropriations! If we don’t advocate for funding the programs dealing with “our” pests, they will shrink.
Watkins and Davidson also worry that whoever is the next secretary of USDA might not support the agency when it seeks to withdraw funds to cover emergencies from the Commodity Credit Corporation – as Secretary Vilsack has.
APHIS and the DHS Customs and Border Protection (CBP) both praised a recent regulatory action that increases user fees for importers having goods cleared at ports. Kevin Harriger, CPB official in charge of agriculture programs, said the new funds would allow CBP to hire 700 new agricultural inspectors (currently there are 2,800 agricultural officials). That sounds great, but … when trade and passenger volumes crashed early in the COVID pandemic, things looked dicey for a while. Plus – as I have argued repeatedly – real protection against pest introductions will come from stronger policies, not ramped-up inspections.
Pathologist Bruce Moltzan reported on the USFS Forest Health Protection program. He pointed out that the USFS has a very limited toolbox. In this fiscal year, the program has about $48 million, after salaries, to support its activities. Projects targetting insects receive 70% of the funding; those targetting pathogens 15%.
2) Invasive hornets
Washington State has not found any new nests of the Northern (formerly Asian) giant hornet (Vespa mandarinia). Miraculous!
However, Georgia detected another species, the yellow-legged hornet (Vespa velutina), near Savannah in August 2023. The Georgia Department of Agriculture, APHIS, and the University of Georgia are working to find nests – which are located at the top of tall pine trees in residential areas. Five nests were found in 2023; another four so far in 2024. Georgia hopes to place traps 100 miles out from each detection site. Like the northern hornet, V. velutina preys on honey bees. It was probably transported by ship or with its cargo.
A third species, V. tropica, has been introduced on Guam.
3) Better Federal-State Cooperation — Sometimes
APHIS and the state phytosanitary officials have set up structures – e.g., Strategic Alliance/Strategic Initiative, or SASI – to work together more closely. CBP joins the coordinating meetings. One program described at the meeting is the effort to contain spread of the box tree moth (Cydalima perspectalis). This effort came out of discussions at last year’s Plant Board meeting, with follow-up gatherings of APHIS, the states, and the nursery industry. The moth is known to be present in New York, Massachusetts, Michigan, Ohio, and now Delaware – plus several Canadian provinces.
A second project concerns how much data to share about state detections of pests – which are recorded in the National Plant diagnostic Network database. These data have accrued over 20 years … and are sought by both other states and academic researchers. States are often reluctant to allow public review of detection data because they fear it will cause other states or private parties to avoid buying plants or other goods from the infested area. While the project team has not yet decided how to deal with these conflicts, they said they were more inclined to share “nonconsequential data” – meaning data on a pest everyone already knows is present, not a pest under regulation or a new detection. In other words, “consequential” seems to pertain to industry profits, not damage to agricultural or natural resources.
EAB-killed ash along Mattawoman Creek, Maryland. Photo by Leslie A. Brice
Craig Kellogg, APHIS’ chief plant health representative in Michigan, reviewed 20 years of dealing with the emerald ash borer (EAB). He is optimistic about the impact of the biocontrol agents that have now been released in 32 states and four provinces. The larval parasitoids are dispersing and EAB densities are coming down. He conceded that over-story and mature ash are still dying, but says ash in long-infested areas are regenerating well. Scientists agree (see Wilson et al. 2024; full citation at end of the blog). Woodpeckers are still the most effective biocontrol agent of EAB for over-story ash, especially in locations where introduced parasitoids are not established. So far, the growing numbers of biocontrol agents are still parasitizing too few EAB larvae to prevent decline of over-story ash trees.
APHIS reported on recent detections of flighted spongy moth from Asia on ships coming to U.S. ports. The program covers four closely related species of Lymantria. All have much broader host ranges than Lymantria dispar, plus the females are capable of sustained flight, so they spread more rapidly.
The principal strategy to prevent their introduction is to require ships that call at ports along the Pacific coast in Russia, China, Japan, and North and South Korea to ensure that the ships’ superstructures and cargo are clean. Before arriving at U.S. ports, the ship’s captain must inform CBP where it has called over the last 24 months. Then, CBP conducts an inspection. If CBP inspectors find a small number of egg masses, they remove the eggs and spray pesticide. If the inspectors detect a large number of egg masses, the ship is ordered to leave port, clean itself, and undergo re-inspection before it can return.
Four countries in the Americas – the U.S., Canada, Chile, and Argentina – and also New Zealand have very similar programs.
Detections follow natural changes in population levels in the exporting regions. APHIS’ program leader, Ingrid Asmundsson, reported on an unfortunate coincidence in 2014. A huge moth population outbreak occurred simultaneously with very low fuel prices in Russia. The latter attracted many ships to call there. An even bigger population surge occurred in 2019. Asmundsson expects another high-moth period this year.
flighted spongy moths infesting a ship superstructure
APHIS is working on putting this program on a regulatory foundation; this would allow the agency to be more specific in its requirements and to impose penalties (other than expulsions from ports). I’ll let you know when the proposed rule is published for comment.
6) Regional Reports: Old Pests, New Pests
Representatives of the regional plant boards informed us of their “new pest” or other concerns.
Gary Fish, president of the Eastern Plant Board, mentioned
awareness that laurel wilt is moving into Virginia and maybe farther north.
elm zig-zag sawfly; photo by Gyorgy Csoka via Bugwood
There was a more general discussion of beech leaf disease. What can be done, given that the disease is so widespread that no one is regulating movement of beech. Gary Fish advised outreach and efforts to reach agreement on management approaches. Chris Benemann, of Oregon, suggested informing other states so that they can decide whether to take regulatory action. A representative of CBP urged engaging stakeholders by asking for their help.
Chris Benemann, President of the Western Plant Board, expressed concern about APHIS’ reduced funding for spongy moth detection and control efforts. She also worries about the recently detected population of Phytophthora austrocedriiin an Oregon nursery. The western states are also focused on several longstanding pest problems – grasshoppers, Japanese beetle; and a new beetle from Australia that is attacking almonds, pistachios, and walnuts.
tree infested by hemlock woolly adelgid; photo by F.T. Campbell
Megan Abraham of Indiana reported that members of the Central Plant Board are concerned about
funding decrease for APHIS and USFS efforts to slow the spread of spongy moth and manage Asian longhorned beetle and emerald ash borer;
She noted that nursery stock is increasingly coming from more distant – and cheaper – producers, raising the risk of new pests being introduced.
Finally, Abraham expressed concern about decreased funding at the same time as the need is growing – and asked with whom states should collaborate in order to reverse this trend.
Kenny Naylor of Oklahoma, Vice President of the Southern Plant Board, concurred that funding levels are a major concern. He mentioned specifically the spongy moth Slow the Spread program and eradication of the Asian longhorned beetle outbreak in South Carolina. Another concern is the Georgia hornet outbreak.
7) Phasing Out Post-Entry Quarantine
APHIS and the NPB have agreed to phase out the post-entry quarantine (PEQ) program. A program review revealed several problems, some of which astound me. When examining plants in quarantine the scientists still relying on visual inspection! And they are looking for pests identified 45 years ago (1980)! While I think PEQ programs can be valuable in preventing introduction of disease agents, as implemented in recent decades it does not. Twenty years ago, citrus longhorned beetles escaped from a “quarantine” area in a commercial nursery in Washington state. These Cerambycids are more than an inch long!
citrus longhorned beetle; photo by Art Wagner, USDA via Bugwood
Part of this phase-out is to transfer plant species harboring pests of concern to the Not Authorized for Importation Pending Pest Risk Assessment (NAPPRA). While the APHIS speaker said that NAPPRA allows the agency to act quickly when it detects evidence of pest risk, I have found lengthy delays. The third round of proposals was published in December 2019! The fourth round of species proposed for NAPPRA listing should be published soon; a fifth round is now in draft inside the agency.
8) Christmas Greens – Spreading Pests
Officials from Oregon, Maine, and Illinois described their concerns about pests being spread by shipments of various forest or plant products, especially Christmas greens. Mentioned were spongy moths, link hemlock woolly adelgid, link elongate hemlock scale, balsam woolly adelgid, link and box wood moth. Part of the challenge is that the vectoring items are often sold by unregulated outlets – multiple stores, Christmas tree lots – and through on-line or catalog outlets. There are also extreme demands on the regulatory enforcement staff during the brief holiday sales season. Several states are unsure whether they have authority over decorative products – although others pointed out that they are regulating the pest, regardless of the object for sale or type of store.
9) Pests in Firewood
Leigh Greenwood of The Nature Conservancy noted that the state agencies that issue firewood regulations – often the plant protection organization (state department of agriculture) — do a good job alerting the public about the risks and rules. However, the public looks first to state parks agencies for information about camping – and those agencies have less robust educational efforts. It is important to put the message where the public can find it when they don’t know it exists – before they include firewood from home in their camping gear.
10) Projects of the North American Plant Protection Organization
The North American Plant Protection Organization (NAPPO) is working on several projects of interest to those of us concerned about tree-killing pests. One project is evaluating risks associated with wood products, especially how well one international regulation, ISPM#15 is working for dunnage. Another projects is testing the efficacy of the heat treatment specified by ISPM#15 (50o C for 30 minutes). A third project — almost completed – is evaluating fumigants that can be alternatives to methyl bromide.
In conclusion, each annual meeting of the National Plant Board is packed with new information, updates on current pests, and comments on by the state agencies who suggest new approached to APHIS and hold the agency to account. It is well worth attending. Information about upcoming meetings of both the national and four regional plant boards is posted on the NPB website, https://www.nationalplantboard.org/
Signatories to the APHIS-NPB strategic alliance
SOURCE
Wilson, C.J., T.R. Petrice, T.M. Poland, and D.G. McCullough. 2024. Tree species richness and ash density have variable effects on emerald ash borer biological control by woodpeckers & parasitoid wasps in post-invasion white ash stands. Environmental Entomology.
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at https://treeimprovement.tennessee.edu/


























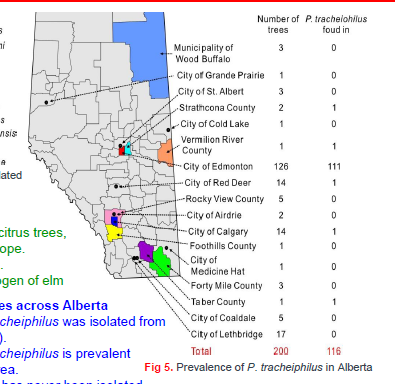























{kind=link}