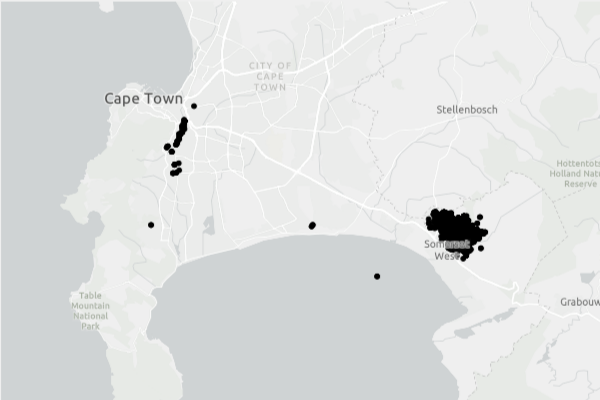
I have learned about the introduction of a North American woodwasp, Sirex obesus, in Brazil. Forestry interests in South America are worried that this woodwasp will cause significant damage to the pine plantations occupying 4.6 million hectares on the continent.
In July 2023, experts at the Estação Experimental de Ciências Florestais at ESALQ/USP in Itatinga, São Paulo, Brazil, investigated dead and symptomatic trees of several Pinus species and subspecies. They expected the causal agent to be Sirex noctilio – a woodwasp native to Europe and North Africa that has caused considerable damage to South American pine plantations since the 1980s (Wilcken et al.).
However, the pine species attacked were not typical hosts for S. noctilio (in Brazil, loblolly pine Pinus taeda). Instead, the infected trees were Caribbean pines, i.e., Pinus caribaea hondurensis, P. caribaea bahamensis, P. caribaea caribaea, P. maximinoi, P. tecunumani. The responsible woodwasp was identified as Sirex obesus. This species is native to the southwestern United States and northern and central Mexico (Wilcken et al.). This species is closely related to S. californicus (Wilcken et al.).
A second outbreak was found in November ~ 130 km away (still in São Paulo state). Scientists have not determined whether the two São Paulo outbreaks are related. Dr. Villacide reports (pers. comm.) that the two populations genetics have been compared, but he does not have the results.
A third population has been detected in a second, neighboring, state, Minas Gerais (Wilcken to Lantschner and Villacide).
Dr. Villacide (pers. comm.) reports that Brazilian scientists are trying to delimit the extent of the outbreaks. Public and private scientists in other countries with pine plantations have begun developing responses.
This is the first record of S. obesus outside of North America (Wilckens et al.).
Little is known yet about this woodwasp’s probable impact. It is clear that it can oviposit in a wide range of pines. In its native range, S. obesus has been reported on three host species: Pinus ponderosa, P. teocote (twisted-leaf pine), and P. leiophylla (no common name; native to Chihuahua – mostly in Mexico, and border areas of New Mexico and Arizona]. In Brazil, as noted, it has been recorded on other species as well as the hybrids P. caribaea x P. elliottii and P. caribaea x P. tecunumanii (Wilcken et al.).
So for purposes of their risk assessment, Lantschner and Villacide assumed that S. obesus can affect any of the species commonly planted in the region: P. taeda, P. elliottii, P. ponderosa, P. contorta, P. caribaea, P. oocarpa, P. patula, P. radiata, and P. tecunumanii (Lantschner and Villacide).
The risk assessment predicts suitable climatic conditions for invasion by S. obesus in 48% of the areas where South American pine plantation occur, particularly in montane and high-altitude regions along the Andean corridor and central-eastern Brazil. Incorporating other factors – host distribution, proximity to invaded areas, and volume of wood imports from Brazil – identified the most vulnerable areas as in southern Brazil, northeast Argentina, the Argentine Patagonia, and central Chile (Lantschner and Villacide).

Preliminary sampling (Wilcken et al.) indicates the impacts could be severe. Mortality varies by species: in the worst cases average mortality approached 43% on P. caribaea hondurensis but only 11% on loblolly pine (P. taeda). They expect mortality rates to increase. Another 30% of P.c. hondurensis trees are dripping resin, a sign of woodwasp oviposition. If these eggs hatch, those larvae will probably kill the affected trees. Such a result would increase total mortality of P.c. hondurensis from 43% to ~ 73%. For P. taeda, the current mortality rate of 11% could rise to 49% as an additional 38% of trees succumb. Following this logic, these areas could experience complete tree mortality within a few years. Given the extent of pine plantations, and possible mortality rates, even a partial spread of S. obesus could lead to significant econ losses.
As second factor is the number of generations per year; the higher the number, the faster woodwasp populations can increase. Wilckens et al. report that adult emergence in Pinus logs maintained in cages indicates that S. obesus could have two or three generations per year.
S. obesus seems to prefer a different climate than S. noctilio. As noted, S. obesus seems to prefer montane and high-altitude climates. S. noctilio is concentrated in lowland temperate and humid regions (Lantschner and Villacide). The newly introduced species might substantially broaden the geographic area where pine plantations might be at risk – although further research is needed to clarify this point.
S. obesus also appears to be spreading at a rapid rate — ~46 km / year. At this rate, Lantschner and Villacide say it could spread throughout all major pine plantation areas in Brazil in less than years.
Sirex woodwasps kill trees by injecting a symbiotic wood decay fungus and a phytotoxic mucus into the tree when ovipositing. The toxin weakens the tree, allowing the fungus to spread, typically killing the tree in as little as three–four months. In North America S. obesus is associated with Amylostereum chailletti. While this species has not yet been confirmed in Brazil, (Wilckens et al.). Brazilian scientists are exploring whether S. obesus might adopt the fungus already present, Amylostereum areolatum, which is associated with S. noctilio.
Two insect species known to feed on woodwasps have emerged from logs infested with S. obesus: Ibalia leucospoides (Hymenoptera: Ibaliidae) and a species of Schlettererius (Hymenoptera: Stephanidae). While these two predators have not proved to be effective controls of woodwasps by themselves, they might become part of a control program. The parasitic nematode, Deladenus siricidicola (Nematoda: Neotylenchidae) used successfully in several South Hemisphere countries to control S. noctilio has not been found in Brazil (Wilckens et al.).
Scientists don’t know the pathway by which S. obesus entered Brazil. Wilckens believes it was via wood packaging; technicians from the Ministry of Agriculture have found some pallets associated with imports that lacked the ISPM#15 mark (Wilckens et al.).
Both Lantschner and Villacide and Wilcken et al. stress the vulnerability of South American pine plantations to introduction of damaging pests. The plantations are reportedly intensively managed, even-aged, regularly spaced monocultures. These conditions can facilitate invasive species establishment and spread by providing abundant host resources and reduced natural enemy pressure. Lantschner and Villacide cite Michael Wingfield that in plantation forestry, introduction of a single pest species can damage large areas of valuable timber.

The family Siricidae contains more than 120 species distributed across the forests of the Northern Hemisphere. In their native ranges they are typically minor or secondary pests (Wilckens et al.). Woodwasps have demonstrated that they can be transported in international commerce – S. noctilio alone has invaded pine stands (native or exotic) in nine countries in Oceania, Africa, and South and North America. Three other species in the family — Urocerus gigas, Urocerus flavicornis and Tremex fuscicornis – have been detected in South America (Wilckens et al.). If each represents a unique threat, countries with widespread pine plantations should enhance their phytosanitary programs. Exporting parties, e.g., the United States and European Union, should assist in efforts to prevent spread of these wood borers. One major step would be to strengthen regulations governing wood packaging material. [To see my criticisms of shortfalls of the ISPM#15 system, scroll down the list of blogs to “Categories” and click on “wood packaging”.]
Lantschner and Villacide cautionthat their assessment is based on a limited record of S. obesus occurrences in its native range. This range might be restricted by factors other than climate, including geographic barriers or biotic interactions (natural enemy pressure or interspecific competition). If so, the species’ potential invasive range might be larger than the climate-based models predict.
Recommendations for management strategies
I applaud Lantschner and Villacide for proposing immediate steps to improve management of the threat posed by introduction of S. obesus. These recommendations should prioritize enhanced phytosanitary inspections of wood products moving between high-risk regions and other South American countries. They suggest that Brazil adopt bilateral agreements with its major trading partners which would specify protocols for woodwaspdetection and quarantines. [Since many of these countries already have established populations of S. noctilio they probably do not have strong phytosanitary measures targeting wood borers at present.] Lantschner and Villacide advise creation of targeted surveillance programs in southern Brazil, northeastern Argentina, Argentine Patagonia, and central Chile. They should focus on sites near major transportation hubs and border crossings. Less intense surveillance should be instituted in regions they classified as medium risk. Again, the focus should be on major points of entry for imported goods and on plantations located near the Brazilian border. They note that preventing spread of S. obesus into new areas will require not only national efforts but also regionally coordinated monitoring, research, and forest health policies.
Lantschner and Villacide also identify priority areas for future research. These include clarifying S. obesus’shost range, the environmental conditions that enable the woodwasp to establish and persist beyond its native range, dispersal rates, and whether S. obesus exhibits pulse-like pop dynamics[long periods of low density interrupted by sudden outbreaks] seen in S. noctilio.
Dr. Villacide (pers. comm.) reports that Brazilian scientists are trying to delimit the extent of the outbreaks. Public and private scientists in other countries with pine plantations have begun developing responses. Dr. Villacide has posted a video from a recent online seminar sponsored by the Southern Cone Forest Health Group. Go to https://youtu.be/uVU6CpFNhlQ?si=lqXtwJTtz5rKXfL3 or
https://sanidadforestalconosur.org/
A wider prespective
Dr. Villacide’s attention to Sirex obesus is part of his broader work on pest issues in South America’s commercial plantations. In another publication (Villacide and Fuetealba 2025; full citation at the end of this blog), he explores how to make these plantations sustainable in the face of rising threats from pests – both introduced and native to the region. Dr. Villacide and Alvaro Fuetealba report that every year 1.2 million hectares of plantations in the Southern Cone are affected by pests. Their vulnerability of will be worsened by the extreme weather events expected under climate change.
These plantations present vast areas of homogeneous stands: ~97% of the Southern Cone planted area consists of exotic tree species – mainly Pinus and Eucalyptus. Typical plantations are high density and managed intensively – including thinning, pruning, and fertilizing – to prompt rapid growth. As Villacide and Fuetealba point out, while these practices maximize wood production efficiency, they also lead to biological homogenization and reduced resilience to pests.
They report that pine plantations are under attack by wood and bark borers that have followed pines to the region, including Sirex noctilio, Orthotomicus erosus, and Cyrtogenius luteus; and now the newly detected Sirex obesus (above). At least two fungal pathogens — Fusarium circinatum and Dothistroma septosporum – have also been introduced. The principal threat to pine plantations from native pests comes from leaf-cutting ants (Atta and Acromyrmex).Eucalyptus plantations are plagued by several insects that have arrived from Australia, including Phoracantha semipunctata, Thaumastocoris peregrinus, and Leptocybe invasa. Pests native to the region that attack Eucalyptus are the Chilean carpenter worm (Chilecomadia valdiviana) and the leaf-cutting ants.

Threat to native conifer
More worrying to me is that introduced pests have entered native forests. Villacide and Fuetealba report that the aphid Cinara cupressi is attacking the native conifer Austrocedrus chilensis. Cordilleran cypress, also called Chilean or Patagonian cedar, is an endemic, monospecific tree in the Cupressaceae family. In southern Argentina and Chile the species forms pure and mixed stands with southern hemisphere beech (Nothofagus spp.) across ~ 160,000 ha. The profile Cinara cupressi on the Global Invasive Species Database is unclear about how many species are in the species complex and their places of origin.
Cordilleran cypress is also under attack by the oomycete Phytophthora austrocedri, an oomycete of unknown origin. This pathogen is of unknown origin. It is now thought to have been present in Argentina since at least the 1960s. P. austrocedri has also been ntroduced to Europe, western Asia, and North America.
Villacide and Fuetealba advocate several actions to might diversify tree species in the plantations to reduce their vulnerability to pests. They note that this recommendation builds on foundational ecological theory, including the resource concentration and natural enemy hypotheses. Diversity-promoting actions should reach beyond any plantation to the landscape level. Managers should consider connectivity of susceptible stands, the number of nutritionally optimal host trees in the landscape, and the availability and quality of hosts in adjacent stands.
Villacide and Fuetealba say mixed plantations can provide additional ecological and economic benefits, such as enhanced stand-level productivity; production of a wider range of commercial and subsistence products; and greater resistance and resilience to natural disturbances, e.g., extreme weather events.
They warn that designing and implementing mixed plantations must reflect ecological interactions and pest dynamics as well as management. There is need for regionally coordinated experimental plantations where scientist could test how variables such as tree species composition, density and spatial arrangement, and silvicultural practices influence pest dynamics, forest productivity, and ecosystem resilience under local conditions. They suggest incorporating sentinel plantings both early-warning systems and decision-support tools at plot and regional scales. Researchers should evaluate pest-specific responses, productivity trade-offs, long-term forest health outcomes under different scenarios.
Since the plantations extend across a multinational region with few natural barriers and uniform silvicultural practices, as well as high levels of trade, so do the pest problems. Therefore, the response must also be regional – e.g., regional experimental plantations and living laboratories. A collaborative approach linking researchers, forest managers, and policymakers is essential to translate experimental findings into practice and develop adaptive, ecol grounded silvicultural strategies. Long-term ecological trials must be embedded in operational contexts and aligned across countries.
SOURCES
Lantschner, V. and J. Villacide. 2025. Invasion Potential of the Recently Established Woodwasp Sirex obesus. Neotropical Entomology. (2025) 54:117 https://doi.org/10.1007/s13744-025-01347-6
Villacide, J. and A. Fuetealba. 2025. Pests in plantations: Challenging traditional productive paradigms in the Southern Cone of America. Forest Ecology and Management 597 (2025) 123127
Wilcken, C.F., T.A. da Mota, C.H. de Oliveir, V.R. de Carvalho, L.A. Benso, J.A. Gabia, S.R.S. Wilcken, E.L. Furtado, N.M. Schiff, M.B. de Camargo, M.F. Ribeiro. 2025. Sirex obesus (Hymenoptera: Siricidae) as invasive pest in pine plantations in Brazil. Scientific Reports. 2025. 15:22522 https://doi.org/10.1038/541598-025-06418-7
Posted by Faith Campbell
We welcome comments that supplement or correct factual information, suggest new approaches, or promote thoughtful consideration. We post comments that disagree with us — but not those we judge to be not civil or inflammatory.
For a detailed discussion of the policies and practices that have allowed these pests to enter and spread – and that do not promote effective restoration strategies – review the Fading Forests report at http://treeimprovement.utk.edu/FadingForests.htm
Or